Коллеги,предлагаю вашему внимаю книгу Н.И. Кривцова "Среднерусские пчелы". Считаю ее лучшим литературным изданием посвященным темной лесной пчеле,при этом нам,коллеги,нужно помнить,что все данные приведенные в этом издании нужно считать усредненными и не абсолютными/эталонными,т.к. они основаны на исследовании пчел взятых с отдельных пасек целых областей/республик,т.е. не была приведена работа по исследованию ста процентов пчелосемей с упомянутых в книге регионов нашей страны. Тем не менее,труд Кривцова информативен,ценен с практической и теоретической стороны вопросов содержания и разведения средне-русских пчел,он должен служить ориентиром для пчеловодов занявшихся породным разведением и содержанием,восстановлением темной лесной пчелы в России (kokafenix).
 СОДЕРЖАНИЕ
СОДЕРЖАНИЕ
Предисловие от редакции
Аборигенные пчелы России
Особенности среднерусских пчел
Популяции среднерусских пчел — исходный материал для селекции
Экстерьерные признаки среднерусских пчел
Хозяйственно полезные признаки пчелиных семей различных популяций
Теоретические основы селекции среднерусских пчел
Селектируемые признаки: наследуемость,изменчивость,повторяемость
Фенотипическая корреляция
Организация, методы и программы селекции пчел
Методы селекционной работы
Селекционные программы, используемые в странах с развитым пчеловодством
Методы селекционной работы в России
Селекция среднерусских пчел
Принципиальная схема чистопородной селекции
Редактор В.В.Александрова
© Н.И.Кривцов, 1995
Предисловие от редакции
В основу этой книги положены многолетние научные исследования, проведенные НИИ пчеловодства (а ранее Всесоюзным НИИ пчеловодства), с целью изучения пчел среднерусской породы и их популяций.
Автор — директор НИИ пчеловодства, доктор сельскохозяйственных наук Н.И.Кривцов — приводит богатейший материал селекционной работы с пчелами среднерусской породы, а также убедительно демонстрирует ее преимущества. Особенно перспективно разведение среднерусских пчел в центральных и северных регионах России, где предпочтение отдавалось завозным южным породам пчел, которые плохо приспособлены для климатических и других условий этих территорий.
Автор искренне благодарит своих коллег — Г.С.Балакиреву, А.В.Седых, T.Н.Оринича, В.Н.Власова, И.Я.Джулая, В.М.Виноградову и М.Д.Большакову за помощь в работе и надеется, что книга будет полезна как ученым-селекционерам, так и пчеловодам-любителям.
АБОРИГЕННЫЕ ПЧЕЛЫ РОССИИ
Особенности среднерусских пчел
Пчелы среднерусской породы (среднеевропейские, темные лесные, собственно медоносные — Apis mellifera mellifera) заселяли отросшие в послеледниковое время леса, адаптировались к природно-климатическим условиям и ко времени появления в западной и центральной Европе человека были аборигенами лесистых равнин. Первоначально пчелы распространились севернее и западнее Альп (Швейцария, Франция, Германия, Центральная Россия). Естественное расселение среднерусских пчел дошло, по-видимому, до Урала.
Большие лесные массивы благоприятствовали пчеловодству. Древнегреческий историк Геродот отмечал, что земли наших предков когда-то были совершенно непроходимы из-за обилия в лесах пчел.
Побывавший в русских лесах Иовий Новокамский писал, что «самая верная жатва получается от воску и меду, ибо вся страна преисполнена плодовитыми пчелами. По лесам и весьма густым рощам здесь можно неоднократно видеть превосходные рои висящих на деревьях пчел, здесь часто находят огромные количества сотов, скрытых в деревах, а в удивительной толщины древесных пнях находят иногда превеликие озера меду» (по А.Морозову, 1978).
Н.М.Витвицкий утверждал, что бортничество и бортевое пчеловодство были некогда для нашего народа тем, чем сейчас является хлебопашество. «Мед и воск были золотом прадедов и дедов наших»,— писал он. В период расцвета бортевое пчеловодство, базировавшееся преимущественно (если не полностью) на среднерусских пчелах, насчитывало в своих угодьях сотни миллионов пчелиных семей, дававших ежегодно почти по 9 млн пудов воска (147,42 млн. кг) и по 600 млн пудов меда (982,8 млн. кг).
Среднерусские пчелы не потеряли своего значения и в настоящее время. Эта порода была рекомендована для разведения в 63 регионах страны, причем в 19 из них, преимущественно северных, она оказалась единственно возможной. Эволюция среднерусских пчел проходила в суровых природно-климатических условиях, в результате чего в процессе приспособления к этим условиям у них появились определенные особенности, закрепившиеся в процессе естественного отбора и отличающие их от пчел других пород. Одним из характерных признаков среднерусских пчел следует считать их поведение при осмотре гнезда. Среднерусские пчелы или «сплывают» к нижнему бруску рамки и повисают гроздьями, или ведут себя слишком возбужденно, быстро перемещаются по соту, взлетают, что очень затрудняет работу с ними. Наряду с этим среднерусские пчелы отличаются сильной злобливостью, и это также является их отрицательным качеством. В периоды отсутствия медосбора даже такие средства защиты от ужалений, как халат, лицевая сетка, обработка дымом, часто оказываются неэффективными.
Большую агрессивность среднерусских пчел по сравнению с кавказскими отмечал, в частности, последовательный сторонник их разведения профессор Г.А.Кожевников (1929). Но он считал, что не все среднерусские пчелы одинаковы в этом отношении.
Ю.В.Страйгис (1974) и Е.М.Петров (1983) указывали на то, что среди местных среднерусских пчел Литвы и Башкирии выделяются семьи повышенной злобливости, которые вместе с тем лучше поддерживают чистоту в улье, более активны и продуктивны.
К сожалению, до сих пор не было предпринято попыток селекции среднерусских пчел на незлобливость, хотя Ф.Руттнер (1968) считал, что такая работа может быть более эффективной, чем отбор по сложным хозяйственно полезным признакам.
Пчелы среднерусской породы печатают мед светлой, «сухой» печаткой, оставляя между восковой крышечкой и поверхностью меда пространство, заполненное воздухом. Это также важный признак породы. У серых горных кавказских и желтых кавказских пчел печатка меда темная, «мокрая».
Для семей среднерусских пчел характерна также нетерпимость маток по отношению друг к другу, в то время как в семьях южных пород нередко наблюдается длительное сожительство в улье двух маток.
Среднерусские пчелы в отличие от пчел южных пород к воровству мало склонны и хуже защищают свое гнездо от пчел-воровок. Возможно, что меньшая склонность к воровству объясняется их слабой предприимчивостью в отыскании источников медосбора. Слабую склонность к воровству следует считать положительным качеством, так как воровитые пчелы затрудняют работу пчеловода на пасеке, а во время подкормок полностью «ограбляют» слабые семьи.
Многие исследователи отмечают у среднерусских пчел высокую склонность к роению. В отдельные годы на пасеках среднерусских пчел до 50—100% пчелиных семей приходят в роевое состояние, а при затянувшемся слабом поддерживающем медосборе начинается бурное роение. По нашим данным, признаки роевого состояния при отсутствии продуктивного медосбора наблюдались с 17 июня по 13 июля у 30% пчелиных семей полесской и уральской популяций и у 60% татарской. За этот период в пчелиных семьях было заложено в среднем по 10 маточников. С появлением медосбора с гречихи (2,0—2,5 кг в сутки) пчелиные семьи вышли из роевого состояния и переключились на сбор меда. При этом они уничтожили не только роевые мисочки, но и зрелые маточники.
Интересно отметить, что склонны к роению пчелиные семьи со старыми матками (3 года и старше). В год вывода матки и на следующий они практически не приходят в роевое состояние. Таким образом, использование молодых маток может быть хорошим приемом против роения у среднерусских пчел.
Матки среднерусских пчел отличаются хорошей плодовитостью, откладывая при благоприятных условиях до 2000 и более яиц в сутки. Среднесуточная плодовитость маток других пород не превышает у серой горной кавказской 1500 яиц, у карпатской—1800 яиц, у краинской—1400—2000 яиц. И только матки итальянской породы, способные откладывать более 2500 яиц в сутки, превосходят среднерусских по этому признаку.
Особенностью среднерусских пчел является усиленное выращивание расплода при слабом медосборе,когда пчелы южных пород, напротив, ограничивают яйцекладку маток.
Если есть возможность выбирать источник медосбора, среднерусские пчелы предпочитают такие медоносы, как гречиха, липа, и работают на них лучше пчел южных пород (В.П.Великанов, 1927; Н.В.Андреева, 1929).
Неоднократно отмечалось, что среднерусские пчелы хорошо собирают мед только при обильном медосборе и значительно отстают от серых горных кавказских при слабом полифлерном медосборе (например, клеверно-луговом) (Г.Д.Билаш, 1962; А.Я.Шекшуев, 1965).
Г.Д.Билаш (1966) считает, что у среднерусских пчел хорошо выражена флороспециализация к нектару гречихи, а также существует сильная привязанность к однажды выявленному источнику медосбора, что может обеспечивать при определенных условиях их преимущество над серыми горными кавказскими.
Некоторые исследователи объясняют различия в флоромиграции, пыльцесобирательной деятельности, использовании того или иного типа медосбора условиями, в которых шла эволюция пород пчел. Если южные породы формировались в условиях длительного, ровного и спокойного медосбора — с большим разнообразием растений с яркоокрашенными цветками, то становление среднерусской породы проходило в условиях лесного типа — с более узкой полифлерностью медосбора и менее яркой окраской цветков медоносов. В первом случае связи пчел с растениями были непрочны, старые рефлексы быстрее угасали, легко вырабатывались новые. В последнем случае пчелы (среднерусские) приспосабливались к короткому активному сезону с бурным медосбором, у них вырабатывалась консервативность в отношении флоромиграции (Н.А.Тушмалова, 1958).
Этим различиям могут быть и другие объяснения, но проведенные наблюдения показывают, что среднерусские пчелы редко мигрируют (Ш.О.Гасанов, 1970) и они лучше собирают пыльцу с бледноокрашенных цветков (Р.К.Миронова, 1974).
Среднерусские пчелы отличаются исключительно высокой зимостойкостью, не зная себе равных по этому признаку среди других пород. Кроме тoгo, они устойчивы к ряду заболеваний (нозематоз, падевый токсикоз).
Все это позволяет успешно разводить их в северных районах страны, где безоблетный период продолжается до 6 месяцев. В местах с сильным поздним медосбором среднерусские пчелы собирают значительные запасы меда.
Из других биологических особенностей среднерусской породы следует отметить следующие:
- чистопородные среднерусские пчелы — темноокрашенные, тергиты не имеют признаков желтизны, стерниты — от темно-серого до темно-коричневого цвета; конец брюшка у рабочих особей более тупой, чем у пчел других пород; посадка спокойно сидящей пчелы низкая, приземистая;
- при осмотре гнезд пчелы очень беспокойны и пугливы, сбегают с сотов вниз и свисают на нижней планке осматриваемой рамки рыхлыми, легко срывающимися гроздьями; злобливость отмечена во всех популяциях, особенно у вологодских и уральских пчел, проявление ее зависит от очень многих внешних факторов; окуривание дымом сильно снижает агрессивность пчел, однако злоупотребление дымом также может привести пчелиную семью в ярость (См.Особенности проявления агрессивности средне-русски...) ;
- при слабом поступлении нектара пчелы охотнее удлиняют медовые ячейки в верхних углах сота, чем отстраивают новые на противоположной стороне улочки; восковые перемычки в верхней части сотов и восковые мостики на печатке меда между двумя сотами отсутствуют либо они редкие, тонкие, лепестковой формы; глубина ячеек в расплодной части рамок всюду одинакова, печатка меда преимущественно белая, «сухая», а при очень слабом, неустойчивом медосборе иногда наблюдается переходный тип печатки;
- при слабом медосборе яйцекладка маток пчелами не ограничивается, выращивание расплода усиливается, а при сильном — яйцекладка резко сокращается и энергия пчел переключается на накопление запасов меда.
Еще Э.Бертран (по Л.Лангелю, 1903—1904) говорил: «Мы бы желали дать один хороший совет начинающим пчеловодам, а именно: не слишком быстро увлекаться иностранными породами пчел. Наша простая северная порода превосходна и во всех отношениях лучше всего пригодна для изучения на ней пчеловодства».
Популяции среднерусских пчел — исходный материал для селекции
Итак, пчелы среднерусской породы по целому комплексу признаков отличаются от пчел других пород, но и внутри породы они тоже неоднородны. Занимая огромный ареал, обитая зачастую в резко отличающихся друг от друга природно-климатических условиях, под влиянием естественного отбора и частично при воздействии человека среднерусская порода образовала обособившиеся группы, приспособленные к тем или иным конкретным условиям. Это характерно для всей породы, а не только для ее части, обитающей в пределах нашей страны.
Еще профессор Г.А.Кожевников (1929), ссылаясь на Е.Цандера, писал, что в Германии различали настоящую темную лесную пчелу, мало склонную к роению, и вересковую пчелу, обитающую на Северо-Западе страны и невероятно ройливую. Ф.Бельденспержер (1928) отмечал, что германские лесные пчелы во многом отличаются от французских. X.Н.Абрикосов (1946) сообщал о голландской вересковой темной пчеле, которая была завезена в Америку в 1638г. Она расселилась в дуплах и расщелинах скал. По наблюдениям В.В.Алпатова (1948), темная лесная пчела, завезенная в США из Голландии, отличается свирепым нравом, в то время как в Англии она сравнительно миролюбива.
Доминирующая на территории Франции темная лесная пчела представлена большим количеством экотипов, приспособленным к местным, узко локализованным, экологическим условиям. Но и в новой местности, в несвойственных им условиях, пчелы сохраняют прежний ритм развития. Таким образом, эта приспособленность наследственна (Ф.Руттнер, 1969; Ж.Луво, 1969).
В Польше издавна содержались такие популяции местных пчел, как среднеевропейская, вересковая и лесная (боровая) (/.Woznica, 1972).
М.Gromisz (1982) склонен выделять в пределах Apis mellifera mellifera такие популяции, как иберийская, среднеевропейская, вересковая, лесная, нигра.
Подобные группы пчел различными исследователями называются по-разному — экотипами, популяциями и даже породами. Именуются такие группы чаще всего по географическому принципу.
На территории бывшего СССР выделены некоторые своеобразные группы среднерусских пчел, которые, с известными допущениями и на уровне современных представлений, можно назвать сложившимися популяциями.
Башкирская (бурзянская) бортевая пчела. Обитает в лесах Бурзянского заповедника, сейчас широко известна. Один из корифеев пчеловодной науки профессор Г.А.Кожевников (1931) об этой пчеле писал так: «В бортях Башкирии и Уральской области мы имеем остаток этой первобытной европейской лесной темной пчелы, которая в настоящее время представляет с точки зрения генетики величайшую драгоценность. Ее надо всемерно беречь от метизации и на ней основать массовое разведение коренной среднеевропейской пчелы, которая тысячелетним отбором в жестокой борьбе за существование среди суровой горной природы выработала в себе выносливость и устойчивость».
Бурзянские пчелы отличаются рядом особенностей: средняя масса рабочей пчелы составляет 109 мг, нагрузка медового зобика —74—87 мг, длина хоботка — 6,10 мм. Пчелы выделяют много воска, дневной принос семьей пчел меда с липы может превышать 10 кг, а яйценоскость маток в июне иногда превышает 2500 яиц в сутки. Роение начинается с первой декады июня и обрывается только с наступлением главного медосбора, при этом закладывается 8—14 маточников. Нередко наблюдается «израивание» семей; такие семьи, как правило, не перезимовывают.
Бурзянские пчелы чутко реагируют на резкие изменения погодных условий и дружно возвращаются к жилью. Воровство наблюдается крайне редко. Одна из характерных особенностей этих пчел — сгрызание вощины в бортях.
В течение августа пчелы готовятся к суровой зиме: собирают пыльцу, мед, заклеивают все щели прополисом, уменьшают летки. Зимовка длится полгода, зимняя температура — минус 40—45°С. При первом весеннем облете бортевые пчелы уже несут пыльцу (В.Г.Генрих, В.А.Тюльпанова, 1958; A.Л.Дулькин, Г.Ф.Трескова, 1953; Е.М.Петров, 1983) (См. Башкирская пчела. , Бурзянские бортевые пчелы и варроатоз. , Заповедник Шульган - Таш и бурзянская бортевая пче...).
Уральская горно-таежная пчела. Исключительно хорошо приспособлена к условиям Северного Урала. Масса рабочей пчелы этой популяции составляет 88— 106 мг, восковая железа развита хорошо, нагрузка медового зобика — 51—72 мг, матка может откладывать до 1800 яиц в сутки. В роевом состоянии закладывается от двух до 28 маточников, чаще 5. Первые вылеты пчелы способны делать в конце февраля — начале марта, уже при температуре плюс 3—4°С. Пчелы могут летать в пасмурную, дождливую и ветреную погоду. Рабочий день на главном медосборе продолжается до 16 ч (В.Синцов, 1951; A.Л.Дулькин, 1958). (См. Средне-русская пчела на севере Урала.).
Алтайская пчела. С.Г.Миньков и другие (1969) приводят данные о появлении пчел в Сибири. Впервые на Алтай (в Усть-Каменогорскую крепость) 30 колод с пчелами были завезены в 1776 г. из Башкирии по ходатайству главного медика пограничных сибирских войск Беренса. Пчелиные семьи хорошо развились, дали по 2—3 роя и по 8 кг меда. Однако зимних кормов было оставлено в колодах недостаточно, и в период зимовки 1778 г. пчелы полностью отошли.
В 1786 г. по просьбе полковника Аршеневского пчелы были повторно завезены из Полесья Киевской губернии. Колоды доставлялись на подводах, и из 24 колод в 12 пчелы были живы. Условия оказались благоприятными для пчел: их стали разводить, а затем и продавать.
За пчелами приезжали отовсюду, и в скором времени вся Чулымская тайга, пространство староказацкой линии между Кузнецком и Бийском, стали базой развития пчеловодства в Сибири. Пчелы определенным образом приспособились к местным условиям, и Н.Ф.Крахотин (1960) отмечает у них ряд особенностей.
Алтайские пчелы не очень ройливы, но в отдельные годы в роевое состояние приходит до 90% семей, при этом закладывается от 5 до 16 маточников. Период роения длится 50—55 дней. Длина хоботка рабочих пчел составляет 6,2 мм, нагрузка медового зобика — 31,5 мг, максимум яйценоскости маток—1575 яиц в сутки. Одна из характерных особенностей этих пчел — встречающиеся при запечатывании меда на гранях крышечек выпуклости в виде жилок светло-желтого цвета, напоминающие по виду завитки шерсти.
Существуют и другие популяции, о которых упоминается в литературе,— алтайская горно-таежная (А.Л.Дулькин, 1960; Н.М.Ершов, 1970), березинская и полесская (В.М.Неручев, 1968; А.С.Гурченко, 1981) и др. Сведения о популяциях среднерусских пчел в Татарии сообщает Т.С.Жданова (1970), а в Мордовии - Т.А.Анциферова и Л.С.Швецова (1970).
С целью изучения генофонда среднерусской породы в 1970—1972 гг НИИ пчеловодства было проведено экспедиционное обследование наиболее типичных областей ее естественного ареала. Цель его — поиск и завоз сохранившихся в чистоте среднерусских пчел, а также выделение внутрипородных популяций и использование перспективного исходного материала для дальнейшей селекции и репродукции.
Располагая крайне скудной информацией по этому вопросу, мы сочли целесообразным подобрать и завезти пчел из мест с наибольшей географической удаленностью друг от друга. Обследовали пасеки, на которые в течение послевоенного периода не завозили пчелиных семей и маток других пород. Отбирая в полевых условиях пчелиные семьи, учитывали следующие признаки: окраску тела пчел и форму конца брюшка, позу пчел в состоянии покоя, активность защиты гнезда, реакцию на дым и поведение на вынутой из улья соторамке, характер отстройки сотов и форму их медовой и расплодной части, печатку меда, наличие и особенности восковых перемычек между сотами. Несмотря на условность каждого из перечисленных признаков, применение их в комплексе обеспечило надежный отбор чистопородного материала, что подтвердилось дальнейшим изучением экстерьера пчел. В результате этой работы были завезены 7 популяционных групп среднерусских пчел, 6 из которых (из Татарии, Новосибирской, Вологодской,Владимирской, Челябинской и Орловской областей) были поставлены на испытание по комплексу хозяйственно полезных признаков.
Экстерьерные признаки среднерусских пчел
Начало морфологическим исследованиям пчел было положено Г.А.Кожевниковым (1900). А.С.Михайлов (1924), В.В.Алпатов (1925) установили зависимость размеров хитиновых частей пчел, и прежде всего длины хоботка, от географической широты местности. В настоящее время экстерьерные характеристики наряду с окраской тела пчел широко применяют для идентификации отдельных географических пород внутри вида. Но выделение внутри вида только географических пород явно недостаточно.
В последние годы значительно возрос интерес к изучению более мелких таксономических единиц пчелы медоносной. Это связано с проблемой сохранения и необходимостью более интенсивного привлечения к использованию в селекционных целях локальных внутрипородных популяций пчел, обладающих ценным генофондом.
Однако выделение локальных популяций вызывает у зоологов определенные трудности. Э.Майр (1968, 1971) отмечает, что степень различий между популяциями бывает от почти полной идентичности до различий практически видового уровня. Границы изменчивости соседних популяций обычно перекрываются. Соседние популяции оказываются сходными в отношении одних признаков и различными в отношении других. Тем не менее локальные популяции слегка отличаются друг от друга генетически и фенотипически. Для установления этих различий обычно требуются очень точные измерения и статистические методы. Если локальные популяции пчелы медоносной, значительно удаленные друг от друга по широте, удается сравнительно легко различать по морфологическим признакам, то подобный анализ популяций из мест, мало различающихся по широте, затруднен. Поэтому перспективным методом, позволяющим анализировать как сходство, так и различие популяций, кроме морфологического анализа может быть сравнение изменчивости признаков. При этом сравнивают как абсолютные значения коэффициентов вариации (С0%) или среднеквадратичных отклонений (сг), так и построенные на их основе графики. Степень изменчивости признаков может, по современным представлениям, свидетельствовать о самых начальных сдвигах в изменении морфологии популяций. В некоторых работах на разных зоологических объектах показана возможность использования параметров изменчивости для изучения популяционной структуры вида (А.В.Яблоков, 1966; А.П.Разоренова, 1975; Ю.И.Новоженов, 1978).
Пчел, завезенных из Владимирской, Вологодской, Орловской, Новосибирской, Челябинской областей и Татарии, мы проанализировали по экстерьеру (табл. 1).
 Между новосибирскими и челябинскими пчелами различий в длине хоботка не установлено. Наибольшие же расхождения по этому признаку (0,15 мм) обнаружены у пчел этих групп по сравнению с орловскими пчелами.
Между новосибирскими и челябинскими пчелами различий в длине хоботка не установлено. Наибольшие же расхождения по этому признаку (0,15 мм) обнаружены у пчел этих групп по сравнению с орловскими пчелами.
Наибольшее значение средней длины крыла отмечено у владимирских пчел, а наименьшее — у новосибирских и челябинских (как и в предыдущем случае). Колебания размеров по этому признаку у всех сравниваемых групп пчел очень близки.
Наименьшая ширина крыла была у челябинских пчел, в остальных группах значения этого признака практически одинаковы и различия не превышают 0,02 мм. Колебания ширины крыла в пределах групп составляют 0,1—0,3 мм.


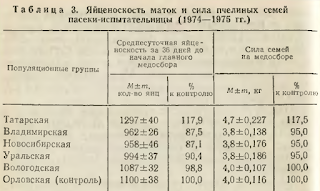 Наивысшую среднесуточную яйценоскость за три учета, предшествующих началу главного медосбора, развивали матки татарской популяционной группы, превосходившие контроль на 17,9%, на втором месте были пчелиные матки местной (орловской) популяционной группы, на третьем — матки вологодской популяционной группы. Максимума яйцекладки матки достигали обычно к концу июня. В этот период среднесуточная яйценоскость лучших маток доходила до 2000 яиц.
Наивысшую среднесуточную яйценоскость за три учета, предшествующих началу главного медосбора, развивали матки татарской популяционной группы, превосходившие контроль на 17,9%, на втором месте были пчелиные матки местной (орловской) популяционной группы, на третьем — матки вологодской популяционной группы. Максимума яйцекладки матки достигали обычно к концу июня. В этот период среднесуточная яйценоскость лучших маток доходила до 2000 яиц.
Яйценоскость маток определяет такой важнейший хозяйственно полезный признак пчелиных семей, как их сила.
Пчелиные семьи татарской популяции по силе на медосборе превышали контроль на 17,5%, на втором месте по этому признаку стояли пчелиные семьи местной и вологодской популяционных групп. Владимирская, новосибирская и уральская популяционные группы были равными по силе и на 0,2 кг уступали контролю.
Различные темпы развития пчелиных семей опытных групп, разная сила на медосборе в значительной мере способствовали их различной продуктивности (табл. 4).

Крайне неблагоприятные погодные условия сезонов 1974—1975 гг. привели к низким медосборам в целом по Орловской области. Так, в Орловском и Урицком районах (в местах расположения кочевых точков пасеки-испытательницы) средний медосбор общественных пасек в 1975 г. составил, соответственно, 14,4 и 13,4 кг
Медопродуктивность пасеки-испытательницы в целом оказалась выше. Наилучшими показателями отличались семьи татарской популяционной группы, на 25,7% по меду и на 13,6% по воску превзошедшие контроль. Превосходство продуктивности татарских пчел над остальными опытными группами было еще большим. Второе и третье места разделили по этому признаку пчелиные семьи вологодской и местной популяционных групп.
Сравнительное испытание шести популяционных групп среднерусских пчел в условиях Орловской области позволило установить некоторые различия между ними. Наилучшей зимостойкостью и наиболее высокой яйценоскостью маток, силой и продуктивностью пчелиных семей отличается татарская популяционная группа, на втором месте оказалась местная (орловская) популяционная группа пчел и на третьем — вологодская.
Во всех изученных популяционных группах среднерусских пчел наблюдается значительная фенотипическая изменчивость хозяйственно полезных признаков,которая может быть предпосылкой эффективной селекции по ним.
Приведенные данные конечно же не отражают все популяции и специфические группировки среднерусских пчел, но они иллюстрируют определенные особенности этих групп и свидетельствуют о значительных генетических резервах среднерусской породы. Учитывая особенности среднерусских пчел, селекционная программа по улучшению породы должна поддерживать имеющийся высокий уровень зимостойкости и развивать потенциальные возможности эффективного использования бурного, но короткого медосбора средней полосы.
ТЕОРЕТИЧЕСКИЕ ОСНОВЫ СЕЛЕКЦИИ СРЕДНЕРУССКИХ ПЧЕЛ
Селектируемые признаки: наследуемость, изменчивость, повторяемость
Подавляющее большинство хозяйственно полезных признаков сельскохозяйственных животных (в том числе и пчел) относится к категории количественных, они контролируются многими генами и детерминируются факторами внешней среды. Наследование полигенных признаков и другие генетико-популяционные параметры современная генетика изучает не на отдельных особях, а в популяциях, с помощью различных математико-статистических методов.
Для генетической характеристики популяций используют такие параметры популяционной генетики, как их фенотипическая и генотипическая изменчивость, наследуемость, повторяемость, фенотипическая и генотипическая корреляция.
По мнению Ф.Ф.Эйснера (1981), эффективность селекции обусловлена величиной селекционного дифференциала, степенью наследуемости селектируемых признаков, быстротой смены поколений, числом признаков, по которым ведут селекцию, характером коррелятивных связей между ними.
Наследуемость позволяет прежде всего судить о генотипическом разнообразии признака, а также дает возможность прогноза селекционной ценности особи по ее фенотипическим проявлениям уже на первых этапах селекционного процесса.
Показатели наследуемости (h2-H2) служат одним из параметров при перспективном планировании селекции. В то же время в каждом массиве животных необходим самостоятельный анализ коэффициентов наследуемости (В.Н.Новоставский, 1969).
Показатель наследуемости (Л2*//2) позволяет точнее установить степень сходства между племенными животными и их потомством, что и определяет его практическое значение в селекции. При более высоком значении h2 отбор по фенотипу лучших особей создает более широкие возможности получения лучшего потомства, а при низких значениях № отбор малоэффективен.
Первые расчеты показателя наследуемости в пчеловодстве касались медопродуктивности (F.Pirchner, F.Ruttner, Н.Ruttner, 1960). Было показано, что № медопродуктивности составляет в среднем 0,2—0,3, что соответствует значениям такового для продуктивности других сельскохозяйственных животных.
Затем исследователи стали принимать попытки использования показателя наследуемости для прогноза селекции на одно поколение (В.Веселый, Р.Шиллер, 1963; М.А.Ел-Бамби, 1965; М.Gromisz, 1972; 1978).
Zawilski (1972) определил коэффициенты наследуемости некоторых экстерьерных признаков пчел. Они оказались выше, чем для показателей продуктивности (h2 = 0,40 — 0,75).
Т.Е.Rinderer, A.М.Collins, М.A.Brown (1983) в Батон-Руже (США) определили h2 устойчивости к нозематозу, продолжительности жизни, реакции на феромон тревоги изопентилацетат. Показатели наследуемости продолжительности жизни пчел и реакции на изопентилацетат составили 0,30—0,32, остальные коэффициенты были недостоверны.
R.F.Moritz (1985) в ФРГ определил коэффициенты наследуемости стадии печатного расплода (h2 = 0,8) и попытался связать продолжительность стадии печатного расплода с устойчивостью к варроатозу. Показатель наследуемости продолжительности жизни пчел у Е.P.Milne (1985) оказался значительно ниже (h2 = 0,196).
Этот же автор изучил наследуемость и целого ряда других признаков; кормодобывающего поведения (на основе лабораторных тестов), массы куколки и т. д.
Приведенные выше сведения показывают, что проблеме наследуемости признаков пчел во всем мире уделяется серьезное внимание. Однако величины показателей наследуемости одних и тех же признаков у разных авторов не совпадают. Это объясняется различными способами расчета показателя h2, а также влиянием конкретных условий. Одна из особенностей h2 заключается в том, что его значение изменяется с изменением условий, зависит от уровня и продолжительности селекционной работы и т. д.
Таким образом, анализ наследуемости признаков необходимо проводить в каждом конкретном случае самостоятельно одним из апробированных методов.
Если наследуемость имеет место при передаче генетической информации от поколения к поколению, то повторяемость (г, г2), по определению Н.А.Плохинского (1969), возникает при реализации одной и той же групповой генетической информации в разных возрастах и в разных условиях: лучшие в раннем возрасте не всегда остаются лучшими в последующих возрастах, лучшие в одних условиях жизни могут стать средними и даже худшими в той же группе, перешедшей в другие условия. Таким образом, повторяемость — это большее или меньшее постоянство рангов особей по их фенотипу в одной и той же группе, но в разных возрастах и в разных условиях жизни.
При высокой повторяемости признака становится эффективным и отбор по нему в раннем возрасте или в любых условиях. В определенном смысле показатель повторяемости служит показателем эффективности раннего отбора по фенотипу или отбора в любых условиях.
Как в начале селекционного процесса, так и в различных его стадиях необходимо иметь данные об изменчивости селективного признака. Чаще всего используется фенотипический коэффициент вариации (Vp)t однако более информативен генотипический коэффициент вариации (Vg), вычисляемый в результате дисперсионного анализа. К сожалению, пока что этот коэффициент селекционерами используется крайне редко (В.А.Мошкин, И.К.Сачли, 1977).
В.П.Коваленко (1976) сделал попытку использовать для выбора методов селекции и оценки ее эффективности на примере яйценоскости кур соотношение показателей изменчивости, наследуемости и продуктивности. Н.А.Соболев (1972) предложил применять в селекционной практике такие параметры, как относительная наследственная стабильность (g2) и вероятность отбора генотипов по фенотипу (р).
В пчеловодстве в той или иной мере используются лишь данные о степени наследуемости признаков и их фенотипической изменчивости. К тому же эти параметры вычисляются различными способами, что приводит к разногласиям между отдельными авторами. Поэтому одной из задач, стоявших перед нами, было подобрать подходящие методы для расчета этих параметров с целью их возможного использования в селекции.
Данные по некоторым генетико-популяционным параметрам среднерусских пчел приведены в табл. 5.
 Все хозяйственно полезные признаки, как видно из значений фенотипических коэффициентов вариации, отличаются значительной фенотипической изменчивостью. Особенно сильно варьируют отдельные показатели зимостойкости (Vp = 50,0—85,6%).
Все хозяйственно полезные признаки, как видно из значений фенотипических коэффициентов вариации, отличаются значительной фенотипической изменчивостью. Особенно сильно варьируют отдельные показатели зимостойкости (Vp = 50,0—85,6%).
Коэффициенты фенотипической вариации за разные годы по одним и тем же признакам отличаются незначительно.
Коэффициент генотипической вариации (Vg) используется селекционерами редко, однако он более информативен, так как позволяет судить о гетерогенности по данному признаку и возможностях отбора в популяции. По всем изученным признакам в среднем Vg меньше Vp примерно в 3,4 раза. Нулевых значений Vg, свидетельствующих об отсутствии возможностей отбора, не зафиксировано ни по одному признаку. Колебания Vg по годам объясняются, очевидно, усилением взаимодействия генотипа и среды в неблагоприятные по погодным и медосборным условиям сезоны.
Значения коэффициентов наследуемости, рассчитанные двумя способами, практически не различались. Разница в сторону снижения коэффициентов наследуемости, рассчитанных с разложением факториального среднего квадрата, наблюдалась лишь при ничтожных «0,1 значениях Н2.
Сравнительно более высокие коэффициенты наследуемости отмечены для максимальной яйценоскости маток и для зимостойкости, выраженной суммой баллов. В первом случае превышение Н2 максимальной яйценоскости над среднесуточной можно объяснить тем, что в период проявления максимальной яйценоскости (конец июня) создаются наиболее благоприятные условия для жизнедеятельности пчелиных семей и работы маток. Во втором случае зимостойкость, выраженная суммой баллов, содержит гораздо более полную информацию о способности пчелиных семей переносить неблагоприятные зимние условия, чем отдельные показатели (отход пчел за зиму, количество печатного расплода весной, активность каталазы ректальных желез), и этот коэффициент наследуемости (Я2) тоже выше.
Значения коэффициентов наследуемости тем точнее чем больше имеется информации об изучаемом признаке. Коэффициенты наследуемости признаков, рассчитанные по усредненным за два года данным, выше рассчитанных по однолетним данным. Так, Н2 медопродуктивности при таком расчете составляет 0,27, среднесуточной яйценоскости маток— 0,24, максимальной яйценоскости 0,31.
Нестабильность значений Н2 может зависеть от многих причин, и в том числе от факторов среды, уровня продуктивности и возраста пчелиных маток (например, в 1972 г. матки были однолетними, а в 1973 г. — двухлетними) .
Высокие значения коэффициентов наследуемости указывают на эффективность отбора по селективным признакам. Однако Н.А.Соболев считает, что, получив низкие значения показателей наследуемости, нельзя считать бесполезным отбор по данному признаку, так как возможна высокая относительная наследственная стабильность признака (g2), если уровень общей вариации невысок. За исключением отдельных показателей зимостойкости (отход пчел за зиму, количество печатного расплода во время первого весеннего осмотра), в наших опытах наблюдалась довольно высокая относительная наследственная стабильность хозяйственно полезных признаков пчел (0,898—0,982).
Высокой относительной наследственной стабильности соответствует и обнадеживающая величина вероятности отбора генотипов по фенотипу (Р), позволяющая предполагать, что для большинства хозяйственно полезных признаков на правильный отбор хороших генотипов можно рассчитывать примерно в восьми случаях из десяти отобранных по фенотипу семей. Для количества печатного расплода во время 1-го весеннего осмотра такой отбор малоэффективен (Р = 0,6340—0,6558), а для отхода пчел за зиму нецелесообразен (Р<0,5000).
Результаты этих исследований позволяют считать, что изученные признаки отличаются большой фенотипической изменчивостью. Коэффициенты генотипической изменчивости ниже фенотипических, однако уровень генотипической гетерогенности достаточен для проведения отбора. Наиболее низкие значения коэффициентов наследуемости, а также относительной наследственной стабильности и вероятности отбора генотипов по фенотипу наблюдаются у признаков, характеризующих зимостойкость. При селекции по этим признакам массовый отбор будет, очевидно, неэффективен, и для их улучшения необходим более высокий уровень селекционной работы.
Коэффициенты наследуемости признаков, характеризующих продуктивность и плодовитость пчел, имеют значения на уровне таковых и для других сельскохозяйственных животных. Расчеты показывают, что при селекции по этим признакам из десяти отобранных по фенотипу пчелиных семей мы можем надеяться на правильный отбор восьми хороших генотипов.
Коэффициенты наследуемости таких признаков, как масса маток, длина хоботка, приближаются по своим значениям к единице. Они лишь в ничтожной степени зависят от паратипических факторов, а коэффициент наследуемости такого признака, как склонность к сбору гречишной пыльцы, составляет всего лишь 0,02.
Один из важнейших хозяйственно полезных признаков — среднесуточная яйценоскость маток. За три года повторяемость среднесуточной яйценоскости среднерусских маток составила:
 Доля действия общих для признака
Доля действия общих для признака
факторов в разные периоды сезона составила в первые два года 26 - 29%, а в 1973 г.— 57% за счет выравненных условий последнего сезона. При этом возникает возможность объективной оценки признака по первым единичным учетам.
Как уже было отмечено, генетико-популяционные параметры селективных признаков динамичны. Мы проследили их изменение на протяжении восьми поколений отбора. Повторяемость определяли как коэффициент корреляции по одному признаку за два смежных года:
 В начале селекции уровень генотипического разнообразия давал возможность рассчитывать на определенный эффект отбора по селективным признакам. В процессе селекции с анализируемыми генетико-популяционными параметрами произошли однозначные изменения: уже к F3 резко снижаются значения h2, г2.
В начале селекции уровень генотипического разнообразия давал возможность рассчитывать на определенный эффект отбора по селективным признакам. В процессе селекции с анализируемыми генетико-популяционными параметрами произошли однозначные изменения: уже к F3 резко снижаются значения h2, г2.
Таким образом, генотипическая изменчивость основных селективных признаков пчел имеет низкие или средние значения (6—34%). То же относится и к степени наследуемости этих признаков. Отсюда можно сделать предварительные выводы: на пасеках с низкой медопродуктивностью массовый отбор вряд ли будет достаточно эффективен. Очевидно, более перспективным окажется индивидуальный отбор с проверкой маток по качеству потомства. Если же уровень селективного признака достаточно высок (например, яйценоскость маток), может быть перспективна дальнейшая селекция по линиям.
Поскольку уровень наследуемости, изменчивости, повторяемости изменяется, необходим постоянный контроль этих генетико-популяционных параметров с целью корректировки селекционного процесса.
Фенотипическая корреляция
Основы учения о корреляции заложены еще Ч.Дарвином и Ж.Кювье. Ч.Дарвин (1859) утверждал, что если человек упорно ведет отбор по какому-либо определенному признаку и, следовательно, все более изменяет его, то в силу таинственных законов корреляции ом почти наверняка вызовет изменения и других частей организма.
Ж.Кювье (по И.И.Шмальгаузену, 1969) писал: «Так как все органы животного образуют единую систему, части которой зависят друг от друга и действуют и противодействуют одна по отношению к другой, никакое изменение не может обнаружиться в одной части без того, чтобы не вызвать соответствующего изменения во всех остальных частях».
И.И.Шмальгаузен (1969), возвращаясь к этой проблеме, отмечает, что различные формы корреляции являются выражением целостности организма в его индивидуальном развитии, которая проявляется в регулярных его реакциях на изменения внешней среды, и даже в реакциях на некоторые нарушения во внутреннем его строении. В процессе эволюции происходит очень медленное, но постоянное накопление корреляционных механизмов общего значения.
В настоящее время установлены корреляции различных признаков. Прежде всего, это корреляция между силой пчелиной семьи и ее продуктивностью. Еще А.С.Михайлов (1927), подтверждая старую истину: «В сильных во время взятка семьях — все спасение», установил достоверную и высокую корреляцию между этими признаками (г = 0,74). Достоверной связи продуктивности с весом пчел и некоторыми экстерьерными признаками он не обнаружил.
В. С. Коптев (1965) показал взаимосвязь между силой семей и медосбором (г = 0,55 — 0,84); G.Н.Cale (1968) установил связь между яйцекладкой матки, сбором пыльцы и медосбором (г = 0,50 — 0,81). Подобную зависимость подтверждают К.Давыденко (1970), Ю.А.Субботин (1971), V.Cavoysky (1977), D.Z.Nelsen, N.Е.Gary (1983). U.Smely (1984), основываясь на высокой корреляции медопродуктивности и площади расплода в начале цветения яблони, предложил использовать последний показатель для прогноза медосбора. Высокую корреляцию между медопродуктивностью и количеством расплода подтверждает I.Woyke (1984).
Предпринимаются попытки выявления корреляций между экстерьерными и интерьерными признаками пчел и маток и медопродуктивностью. Еще В.О.Пикель (1926) писал о зависимости медопродуктивности от длины хоботка и размеров тела пчел, а С.Г.Петров (1927) пытался прогнозировать медопродуктивность по развитию большой ядовитой железы. Однако А.Велих (1929) отрицал связь между длиной большой ядовитой железы, злобливостью и медопродуктивностью.
А.Н.Колесников (1959) установил прямую корреляцию между продуктивностью и такими признаками, как длина крыла, количество зацепок, длина и ширина воснового зеркальца и особенно ширина первого членика задней лапки, размеры крыла и тергитов.
I.Heitmanek (1960) отрицал влияние длины хоботка рабочих пчел на медосбор. Вместе с тем М.Н.Виноградов, В.М.Фролова (1969) убедительно показали на примере серых горных кавказских пчел влияние целого ряда экстерьерных признаков, и прежде всего длины хоботка, на медопродуктивность. В.Г.Кашковский (1965) считал, что для Кемеровской области более подходят пчелы с коротким хоботком.
С.P.Milne (1980) выявил связь веса куколок на стадии розовых глаз с медопродуктивностью. Г.А.Аветисян (1961) установил значительную корреляцию между количеством яйцевых трубочек, массой матки и ее плодовитостью, а Тхай-Бин-Ван (1975) — массой маток, количеством яйцевых трубочек и суммой длин 2—5-го тергитов. А.Н.Мельниченко и др. (1968),А.С.Тришина, Н.Д.Шмелева (1974) отрицали наличие какой-либо связи между яйценоскостью маток, количеством и длиной яйцевых трубочек, размерами маточников и массой маток.
Однако многие авторы даже рассчитывали коэффициенты корреляции между интерьерными и экстерьерными признаками. А.С.Михайлов (1927) установил связь между весом рождающейся пчелы и ее размерами, а также между длиной переднего крыла и числом зацепок на заднем крыле у тульских пчел. Т.Szabo (1973) нашел прямую зависимость между весом народившихся маток и их весом после прекращения яйцекладки.
Обнаружена корреляционная зависимость таких признаков, как площадь воскового зеркальца и сумма ширин 3-го и 4-го тергитов (L.Bornus, 1960), длина 3-го тергита и общая длина тела (В.Губин, 1969).
В. В. Тряско (1961), Г.Д.Билаш, В.Т.Желтякова (1965) рассчитали целый комплекс корреляционных связей между экстерьерными признаками.
В.В.Алпатов (1927) установил связь между силой пчелиной семьи и весом выводящихся в ней пчел (г= = 0,489), но А.С.Михайлов (1927) эту зависимость не
подтвердил.
М.В.Жеребкин (1969) выявил довольно высокую корреляцию между активностью инвертазы глоточных желез и медопродуктивностью (г=0,51— 0,87) и предложил использовать этот признак для прогнозирования медопродуктивности.
Т.Szabo (1974) предположил прямую взаимосвязь между массой матки на выходе и в период яйцекладки.
С.Mraz (1981) считает, что агрессивность пчел часто связана с их высокой жизнестойкостью, резистентностью к заболеваниям и высокой медопродуктивностью.
Таким образом, селекционеры предпринимают попытки расчета корреляций между самыми различными признаками пчел. Одна из целей этой работы — облегчить отбор признаков, так как селекция по целому комплексу параметров менее эффективна, но более трудоемка. При установлении же положительной корреляции возможен косвенный отбор одних признаков по другим, более доступным для учета. Изучение корреляций позволяет проводить раннюю оценку, прогнозировать развитие селективных признаков. Правда, мнения специалистов о существовании корреляции между одними и теми же признаками противоречивы, а то и диаметрально противоположны. Но они лишь подтверждают положение о том, что коэффициенты корреляции имеют смысл лишь в определенных условиях и на конкретных объектах, и в каждом отдельном случае их следует рассчитывать самостоятельно.
Селекция пчел по таким признакам, как медопродуктивность и зимостойкость пчелиных семей, яйценоскость маток, как, впрочем, и по большинству хозяйственно полезных признаков других сельскохозяйственных животных, затруднена вследствие того, что эти признаки контролируются многими генами и в значительной степени детерминируются факторами внешней среды.
Относительно высокое влияние этих факторов на коэффициенты наследуемости хозяйственно полезных признаков отмечают многие исследователи независимо от различий в используемых методиках расчетов (Н.А.Плохинский, 1964, П.Рокицкий, 1978).
В данной ситуации нельзя пренебрегать возможностями косвенного отбора пчел на медопродуктивность, зимостойкость, яйценоскость маток, используя коррелятивные связи.
Эффективность селекции, как уже было сказано, зависит как от степени наследуемости признаков, величины селекционного дифференциала, быстроты смены поколений, числа селективных признаков, так и от характера коррелятивных связей между ними.
Изучение коррелятивных связей между признаками пчелиных семей и выявление возможностей использования косвенного отбора в селекции пчел будут способствовать эффекту селекции.
Медопродуктивность является главным селективным признаком большинства селекционных программ, и поэтому изучение коррелятивных связей хозяйственно полезных и экстерьерных признаков непосредственно с медопродуктивностью представляет наибольший интерес.
Ниже приводятся данные, показывающие коррелятивные связи некоторых признаков с медопродуктивностью:
 Анализ этих данных показывает, что экстерьерные признаки рабочих пчел, характеризующие размеры крыла и тела, положительно коррелируют с медопродуктивностью. Хотя колебания размеров этих признаков и не выходили за рамки стандарта породы, пчелиные семьи с относительно большим развитием данных признаков у рабочих пчел отличались и более высокой медопродуктивностью.
Анализ этих данных показывает, что экстерьерные признаки рабочих пчел, характеризующие размеры крыла и тела, положительно коррелируют с медопродуктивностью. Хотя колебания размеров этих признаков и не выходили за рамки стандарта породы, пчелиные семьи с относительно большим развитием данных признаков у рабочих пчел отличались и более высокой медопродуктивностью.
Зимостойкость пчелиных семей также положительно коррелирует с медопродуктивностью, хотя между оценками этих признаков проходит 3—4 месяца — практически весь активный пчеловодный сезон. Следовательно, по результатам зимовки еще ранней весной можно сделать предварительный прогноз медопродуктивности.
Значительной по силе положительной корреляцией связана с медопродуктивностью и среднесуточная яйценоскость маток, а также определяющиеся ею максимальная яйценоскость, общая масса пчел на период медосбора и летная деятельность пчел.
Из признаков, коррелирующих с медопродуктивностью, более подробного рассмотрения заслуживают такие сложные хозяйственно полезные признаки, как зимостойкость пчелиных семей и яйценоскость маток. В зависимости от целей селекции они могут быть и самостоятельными. Зимостойкость пчелиных семей складывается из целого комплекса более простых показателей, находящихся в коррелятивной зависимости (табл. 6).
 Данные табл. 6 показывают, что вследствие положительной корреляции большой расход корма за зиму может привести к нежелательным последствиям — большому отходу пчел за зиму, переполнению кишечника пчел экскрементами (зимующие пчелы при благополучной зимовке не освобождают кишечник до весеннего облета и, как следствие, к большей опоношенности гнезд. Степень опокошенности гнезд коррелирует положительно с зимним отходом пчел, наполненностью кишечника пчел экскрементами и отрицательно — с количеством печатного расплода весной.
Данные табл. 6 показывают, что вследствие положительной корреляции большой расход корма за зиму может привести к нежелательным последствиям — большому отходу пчел за зиму, переполнению кишечника пчел экскрементами (зимующие пчелы при благополучной зимовке не освобождают кишечник до весеннего облета и, как следствие, к большей опоношенности гнезд. Степень опокошенности гнезд коррелирует положительно с зимним отходом пчел, наполненностью кишечника пчел экскрементами и отрицательно — с количеством печатного расплода весной.
Наполненность кишечника пчел экскрементами, положительно коррелирующая с такими показателями, как зимний отход пчел, расход корма за зиму, степень опоношенности гнезд, и отрицательно — с количеством печатного расплода весной, может служить в качестве сигнального признака успеха зимовки. Данные по этому признаку можно получать в результате взвешивания кишечников у пробных групп пчел, взятых еще в зимовнике; можно наблюдать и за динамикой накопления экскрементов в течение зимы
Степень опоношенности гнезд также можно использовать в качестве сигнального признака успеха зимовки, определяя ее как в зимовнике, так и сразу же после выставки при беглом весеннем осмотре пчелиных семей, когда погодные условия не позволяют производить полную разборку гнезд.
Количество печатного расплода при 1-м весеннем учете, отрицательно коррелируя с зимним отходом пчел, расходом корма за зиму, степенью опоношенности гнезд и наполненностью кишечника экскрементами, также может служить косвенным показателем успеха зимовки. Кроме того, этот показатель положительно коррелирует и с общей массой пчел, выращиваемых к медосбору (л=0,32 — 0,65). Было изучено влияние на показатели зимостойкости и некоторых других признаков (таб. 7).
 Пораженность пчел нозематозом, выраженная количеством спор в среднем на одну пчелу каждой семьи, коррелирует положительно с каловой нагрузкой и отрицательно— с количеством печатного расплода весной.
Пораженность пчел нозематозом, выраженная количеством спор в среднем на одну пчелу каждой семьи, коррелирует положительно с каловой нагрузкой и отрицательно— с количеством печатного расплода весной.
Наблюдается тенденция к положительной связи этого признака со степенью опоношенности гнезд.
Активность каталазы ректальных желез пчел коррелирует отрицательно с каловой нагрузкой, расходом корма за зиму и имеет тенденцию к отрицательной связи со степенью опоношенности гнезд. Слабая положительная корреляция наблюдается между активностью каталазы ректальных желез пчел и количеством печатного расплода.
Увеличение процента сахарной добавки в зимних кормах приводит к снижению расхода корма за зиму, к уменьшению степени опоношенности гнезд и величины каловой нагрузки, и в то же время отрицательно сказывается на количестве печатного расплода весной. Отмечая положительное влияние замены части меда в зимних запасах сахаром на некоторые показатели зимостойкости, необходимо в то же время определить оптимальные размеры сахарной добавки применительно к определенным природно-климатическим условиям и конкретным породам пчел.
Зимостойкость пчелиных семей, выраженная комплексным баллом, оказывает определенное влияние непосредственно на медопродуктивность (см. табл. 7) и более выраженно влияет на яйценоскость пчелиных маток:
 Наибольшее влияние результаты зимовки оказывают, естественно, на яйценоскость в раннем периоде развития, отмечаемую при 1-м весеннем учете. Коррелятовные связи в этом случае наиболее тесные.
Наибольшее влияние результаты зимовки оказывают, естественно, на яйценоскость в раннем периоде развития, отмечаемую при 1-м весеннем учете. Коррелятовные связи в этом случае наиболее тесные.
Несколько ниже значения коэффициентов корреляции зимостойкости с яйценоскостью (определяются за 36 дней до начала главного медосбора), и совсем незначительна корреляция зимостойкости с максимальной яйценоскостью непосредственно перед началом главного медосбора.
Максимальная яйценоскость связана сильной положительной корреляцией со среднесуточной яйценоскостыо (г=0,67 — 0,91), что дает основание судить о среднесуточной яйценоскости по значению максимальной. Это обстоятельство дает возможность при крупномасштабной селекции проводить экспресс-оценку общей яйценоскости по показателю максимальной. С максимальной яйценоскостью маток положительно коррелирует их масса (г=0,31), а с массой — длина и ширит 3-го стернита, т. е. размеры тела (г=0,56 — 0,77). Это дает возможность проводить косвенный отбор на яйце носкость по массе и размерам маток сразу же после их вывода.
Таким образом, изученные хозяйственно полезны признаки пчел представляют собой комплекс устойчивых взаимосвязей. Направление коррелятивных связей по годам не изменялось, а их сила колебалась незначительно. Анализ коррелятивных связей показывает возможность проведения косвенного отбора по селективным признакам и осуществление ранней оценки селенкционного материала.
Мы проанализировали также возможности косвенного отбора по качеству маток. Пчеловоды знают, что даже матки с известным происхождением и несомненно хорошей наследственностью не всегда показывают хорошую яйценоскость. Другими словами, ценные гены иногда могут реализоваться в посредственных и также плохих фенотипах. Причиной этому могут быть плохие условия содержания и кормления маток в семьях-воспитательницах, недостаток пчел-кормилиц, технические недоработки при прививке личинок и целый ряд других факторов, которые трудно учесть и устранить.
В этой связи возникает необходимость тщательного отбора получаемых маток по признакам, доступным для непосредственной оценки — внешнему виду, размерам, массе, т е. по фенотипу. Такой отбор повышает также результативность селекционной работы. Здесь вполне уместно известное изречение М.Ф.Иванова: «Хорошие генотипы следует искать среди хороших фенотипов».
Учитывая такую биологическую особенность пчелиных маток, как короткий срок жизни, особое значение приобретают методы ранней оценки маток по косвенным признакам — размерам маточников, массе неплодных маток по выходе из маточников и т. д.
Однако данные о корреляционных связях между экстерьерными и интерьерными признаками пчелиных маток, а также их яйценоскостью весьма противоречивы.
Так, П.М.Комаров (1937), К.Вейс (1972) не установили четкой связи между весом маток и числом яйцевых трубочек. Г.А.Аветисян (1967) обнаружил лишь слабую связь между этими признаками.
Л.А.Хидешели и Н.В.Мчедлишвили (1973), Ф.Таранов (1974) выявили у маток серых горных кавказских пчел, а Хорошилов (1973) у маток дальневосточных пчел высокую положительную корреляцию между весом маток и количеством яйцевых трубочек.
А.Н.Мельниченко и др (1968), А.С.Тришина и Н.Д.Шмелева (1974) провели подобную работу, рассматривая разные породы пчел, и в целом выявили слабую связь между весом маток и количеством яйцевых трубочек. Они нашли положительную связь между числом яйцевых трубочек и объемом маточников у кавказских, краинских и дальневосточных маток. Отмечена высокая положительная корреляция между числом яйцевых трубочек и глубиной маточников у итальянских и дальневосточных маток. В большинстве же случаев не наблюдается достоверной корреляции между числом яйцевых трубочек и размером маточников.
Меньше разногласий вызывает характер корреляции между объемом маточников и весом неплодных маток а также размерами тергитов и стернитов и весом маток. Здесь отмечается положительная корреляция. (Г.А.Аветисян и др., 1971; П.О.Губа, 1973; А.Л.Хидешели и Н.В.Мчедлишвили, 1973).
В результате наших исследований (Н.И.Кривцов, М.Д.Большакова) установлена положительная корреляция между массой маток и большинством экстерьерных признаков (табл. 8, 9). Особенно четкая взаимосвязь наблюдается между массой маток и размерами 3-го стернита.

Количество яйцевых трубочек более тесно связано с шириной 3-го тергита и 3-го стернита, с длиной 3-го тергита корреляции не обнаружено. Не удалось установить и связи массы неплодных маток с количеством яйцевых трубочек. В опыте маток взвешивали по выходе из маточников и каждую нумеровали. Нередко попадались матки, у которых в каждом яичнике насчитывали до 170 яйцевых трубочек, а весили они как 180 так и 210 мг. В то же время по 140 яйцевых трубочек в одном яичнике было, у маток массой 160, 180, 210, 220 мг.
 Небольшую, но достоверную положительную связь наблюдали между массой оставшегося в маточниках корма и массой неплодных маток. Их проанализированных 472 маточников все легкие матки (массой менее 180 мг) вышли из маточников, в которых не оставалоси корма, а среди тяжелых маток только 10% вышли из маточников, где не было неизрасходованного корма. От общего же количества взвешенных маток, вышедших из маточников, где не оставалось корма, легких и тяжелых маток оказалось по 3%.
Небольшую, но достоверную положительную связь наблюдали между массой оставшегося в маточниках корма и массой неплодных маток. Их проанализированных 472 маточников все легкие матки (массой менее 180 мг) вышли из маточников, в которых не оставалоси корма, а среди тяжелых маток только 10% вышли из маточников, где не было неизрасходованного корма. От общего же количества взвешенных маток, вышедших из маточников, где не оставалось корма, легких и тяжелых маток оказалось по 3%.
Обнаружена положительная связь между глубиной маточников и массой неплодных маток, а также между объемом маточников и массой неплодных (г = 0,46) и плодных (г = 0,22) маток. Масса неплодных и плодных маток тесно коррелирует, что открывает возможности объективного раннего отбора по первому признаку.
В опыте 1985 г. масса 51 неплодной матки составила в среднем 206 мг, а масса этих же маток после того , как они стали плодными, достигла 229,5, т. е. увеличилась в среднем на 23,5 мг.
Хотя проследить влияние массы маток на их яйценоскость сложно из-за множества паратипических факторов, определяющихся внешними и внутренними условиями жизни пчелиной семьи, эти признаки положительно коррелируют. В наших исследованиях с максимальной яйценоскостью маток коррелировала их масса (взвешивали неплодных маток, г = 0,31 ±0,174, гс = 32). Отмечена незначительная корреляция массы неплодных маток с силой пчелиных семей на медосборе (г=0,41 rfc ±0,130, п—72; 1985 г.). Приведенные данные также свидетельствуют в пользу косвенного отбора на яйценоскость по массе неплодных маток. Однако если проанализировать влияние массы неплодных маток на максимальную яйценоскость плодных маток в узких пределах (от 195 до 225 мг), то здесь корреляция отсутствуем (г = —0,11, гс = 40; 1985 г.).
На наш взгляд, можно выделить два основных признака, предварительно характеризующих качество маток,— их массу и количество трубочек в яичниках. С учетом изученных корреляций можно составить следующую схему косвенного отбора качественных маток, отбраковка неполноценных маточников; отбор неплодных маток по оптимальной массе (220 мг) и размерам 3-го тергита и стернита, особенно по их ширине (в производственных условиях отбор проводится визуально по размерам тела); контроль за наличием оставшегося корма в маточниках по выходе матки (отсутствие корма показывает неполноценность кормления); контроль за внешним видом маток.
Основная и окончательная оценка качества маток проводится непосредственно по их яйценоскости в пчелиных семьях.
В то же время, следует иметь в виду, что масса плодных маток в пределах, близких к стандарту, может и не коррелировать с их максимальной яйценоскостью и силой пчелиных семей на медосборе (в 1985 г. мы выявили даже тенденцию к отрицательной корреляции между этими признаками). Это дает основание предполагать, что более плодовиты матки с массой, приближающейся к 220 мг (стандарт), а не к 250 мг (в опыте). Следовательно, целесообразно отбирать маток оптимальной массы, бракуя легких (ниже стандарта) и не стремясь к отбору чересчур тяжелых (больше 250 мг).
Таким образом, значения генетико-популяционных параметров основных селективных признаков среднерусских пчел, таких, как фенотипическая и генотипическая изменчивость, наследуемость, повторяемость, дают основание рассчитывать на определенный успех селекции. Их необходимо анализировать на всех стадиях селекционного процесса.
Также должен сопутствовать селекционному процессу анализ корреляций, что позволяет более объективно оценивать селекционные признаки, дает возможность, прогнозирования развития одних из них по другим, а также осуществления косвенного отбора.
ОРГАНИЗАЦИЯ, МЕТОДЫ И ПРОГРАММЫ СЕЛЕКЦИИ ПЧЕЛ
Методы селекционной работы
Медоносные пчелы — насекомые с высоким уровнем общественной жизни, живущие семьями, состоящими из одной самки (матки), нескольких десятков тысяч ее дочерей (рабочих пчел) и нескольких сотен сыновей (трутней), которые появляются в благополучных семьях только летом и выполняют функцию размножения (См. Биология пчелиной семьи. ).
Трутни гаплоидны, т. е. они развиваются из неоплодотворенных яиц, тогда как матки и рабочие пчелы диплоидны — они происходят из яиц, оплодотворенных спермой, хранящейся в сперматеке матки и полученной от нескольких (до 15) трутней. Спаривание маток и трутней происходит в воздухе, иногда на значительном расстоянии от пасеки, что затрудняет его контроль (См. Процесс спаривания. ).
В то время как трутень спаривается один раз в жизни и только с одной маткой, матка чрезвычайно плодовита и может откладывать 2—3 тыс. яиц в сутки. Образно говоря, трутни представляют собой гаметы от матки. Отмеченные особенности показывают целесообразность селекции по женской линии.
Многие хозяйственно полезные признаки медоносной пчелы (медопродуктивность, воскопродуктивность) являются результатом деятельности рабочих пчел семьи, и селективная ценность этих признаков оценивается не через участвующих в спаривании маток и трутней, а через рабочих пчел. Таким образом, пчелиная семья в целом представляет собой своеобразный и оригинальный селекционный объект.
Как и в других отраслях животноводства (конечно, с учетом специфических особенностей), в пчеловодстве используют методы чистопородного разведения и скрещивания.
Подчеркивая творческое значение селекционной работы вообще, известный советский генетик Н.И.Вавилов (1934) писал: «Селекция по существу есть вмешательство человека в формообразование животных и растений; другими словами, селекция представляет собой эволюцию, направляемую волей человека». Формы и методы селекционной работы могут быть различны: одни селекционеры предпочитают селекцию с использованием нескольких пород, другие являются сторонниками чистопородного разведения (См. Старая история о наилучшей пчеле...).
Еще известный швейцарский ученый-пчеловод Крамер (1898—1899), рассуждая о выборе породы, отмечал, что местная пчела, «...которая в течение целых тысячелетий приноравливалась к условиям нашего климата и акклиматизированная подходящими способами, должна уже по одному тому представлять из себя самый благородный продукт для разведения, так как она менее всего склонна к вырождению».
Рассмотрим схемы селекционной работы в некоторых странах с развитым пчеловодством.
Задача племенной работы в пчеловодстве — постоянное улучшение пород пчел (См. Породы пчел.). Следствием такого улучшения является развитие основных хозяйственно полезных признаков (медопродуктивности, воскопродуктивности) и их устойчивая передача из поколения в поколение. Хотя показатели продуктивности пчел складываются из множества факторов (длина хоботка рабочих пчел, нагрузка их медового зобика, продолжительность жизни, работоспособность, предприимчивость в отыскании источников медосбора, устойчивость к заболеваниям, зимостойкость, плодовитость маток и др.) и зависят от них, все же селекцию целесообразно вести не по этим отдельным показателям, а по результатирующему признаку, в частности медопродуктивности.
Чтобы оценка и отбор пчелиных семей были объективными, необходимо их проводить среди возможно большего количества сравниваемых пчелиных семей.
В любом случае первым этапом селекционной работы должно быть получение линий на основе чистопородного разведения, максимальное использование их генетических особенностей для достижения наибольшего хозяйственно полезного эффекта. Пути и формы такой работы могут быть различны. Однако развитие таких линий ограничено, и наступает этап, когда возникает необходимость проведения межлинейного скрещиваиия как внутри породы, так и между породами
F.Kobel (1968), следуя концепции Крамера о преимуществах чистопородного разведения местных пчел, тщательно переработал девятое издание его книги, изменив название на «Чистопородное разведение медоносной пчелы». Это фундаментальный труд, полезный не только в качестве практического руководства по чистопородной селекции для швейцарских пчеловодов, но интересный для всех селекционеров.
Важнейшим моментом Ф.Кобел считает составление тщательного плана селекционной работы. Решающее значение имеет отбор племенных пчелиных семей. При этом следует контролировать также продуктивность семей с матками-сестрами,. тетками и др., т. е. вести отбор по происхождению и по качеству потомства. Средством для проведения такого отбора является балльная оценка. После оценки исходного материала необходимо выбрать не менее шести племенных маток, вывести от них дочерей и сформировать группы семейств. После двухлетнего испытания отбирают лучших.
При разведении по линиям вследствие инбридинга может ослабляться жизнеспособность пчелиных семей. В таком случае встает вопрос об «освежении крови». Хорошая селекционная работа невозможна без использования изолированных случных пунктов. Однако замена маток на соседних пасеках и без надежной изоляции (См. Гаранты чистопородности при репродукции в неизолир...), по мнению Ф.Кобела, может обеспечить в течение нескольких поколений хороший селекционный эффект.
К.Dreher (1976) дал обстоятельную и обобщенную характеристику методам чистопородного разведения и скрещивания в пчеловодстве. Среди предпосылок успешной селекционной работы он выделяет ясность селекционной цели, наивысшую продуктивность при возможно более низких затратах, единообразие состава и надежность наследования ценных признаков. Селекционер может влиять на генотип в результате отбора (накопление плюс-вариантов и исключение минус-вариантов) или используя эффект гетерозиса (См. О гетерозисе и вреде не контролируемой гибридизаци...). При этом следует иметь в виду, что полученные в последнем случае изменения не наследственны. Различные методы внутрипородной селекции можно классифицировать следующим образом.
Инбридинг (инцухт) предполагает: тесное линейное разведение, приводящее через несколько поколений к образованию инбредных линий; широкое линейное разведение; тесное групповое разведение (например, внутри штаммов Тройзен, Скленар); широкое групповое разведение (например, внутри экотипов альпийской карники).
Аутбридинг (аусцухт) предполагает; планомерное 4-линейное скрещивание инбредных линий; планомерное скрещивание различных по происхождению групп одного экотипа; планомерное скрещивание различных экотипов.
Среди методов межпородного скрещивания можно выделить получение и промышленное использование F-1,многократное пользовательное скрещивание, ротационную селекцию, вытеснительное скрещивание, породообразующее скрещивание.
Одним из первых исследование тесного инбридинга у пчел провел современник Дзержона и Берлепша Ф.Фогель, который от двух семей египетских пчел вывел множество поколений. Начиная с 6—7-го поколения пчелиного расплода становилось все меньше, семьи делались нежизнеспособными.
После второй мировой войны Г.Виддель из штамма Скленар путем близкородственного спаривания вывел свой штамм Альба. Он достиг крайнего усиления признаков карники, но при этом также значительно снизилась жизнеспособность пчел. Такая же проблема возникла у доктора Кейла из США, получившего для фирмы «Дадан и сыновья» несколько инбредных линий. Содержание таких линий для последующего кроссирования было сопряжено с большими издержками.
Как только вследствие инбридинга происходит обеднение генов, повышается (до 50%) выпадение расплода пчел. Правда, как уже было сказано выше, против нежелательных последствий инбридинга у медоносных пчел выработался целый ряд приспособлений: спаривание половых партнеров происходит далеко от своей пасеки и в основном с неродственными партнерами; молодые матки за 1—3 брачных вылета спариваются в среднем с 8—9 трутнями; гаплоидность трутней способствует тому, что вредоносные гены у них не могут оставаться в скрытом состоянии, они проявляются внешне (белоглазость, бесфасеточность глаз). Такие трутни не выдерживают конкуренции в местах их сбора.
Нетесный инбридинг при широком линейном разведении (спаривание кузинов и родственников с более отдаленным родством) вполне приемлем для селекций при наличии надежных пунктов спаривания или инструментального осеменения.
Тесное групповое спаривание осуществляется внутри определенной популяции. Пока штаммы не сужаются по линиям, опасности инбридинга нет.
Успешно можно практиковать широкое групповое разведение.
Продукты скрещивания инбредных линий отличаются более высокой продуктивностью за счет эффекта гетерозиса. Однако этот эффект неустойчив, и через несколько поколений продуктивность снижается до уровня исходного материала и ниже. Широко известны гибриды кукурузы, полученные на основе инбредных линий. По этому же принципу были созданы гибридные штаммы пчел Старлайн и Миднайт в США.
Достаточно распространено и планомерное скрещивание внутри экотипа, а также различных экотипов внутри породы. Этот метод также дает возможность получать значительный эффект гетерозиса, но так бывает далеко не всегда, и поэтому в каждом конкретном случае надо проводить специальные исследования.
При межпородной гибридизации, когда используется F-1 ,селекционеры должны для получения помесей брать только чистые породы.
С середины прошлого столетия начался завоз иноземных пород пчел, что повсеместно привело к бесплановой метизации и уничтожению генофонда местных пород См. Экологический гетерозис пчелиных семей и ошибки по...). Содержание чистых пород и плановое получение помесей первого поколения требует предварительных расходов и четкой организации (См. О гетерозисе и вреде не контролируемой гибридизаци...).
В пчеловодстве возможны и многократные межпородные скрещивания. Этот способ успешно применил Адам при создании бакфастовской пчелы. При этом пчелы итальянской породы скрещивались с местными английскими пчелами.
Следует отметить также вытеснительное скрещивание, довольно широко используемое в пчеловодстве. При этом способе в целом хорошая порода, имеющая лишь некоторые недостатки, скрещивается с менее ценной, но не имеющей этих недостатков. Последующие генерации скрещивают снова с предпочитаемой породой. Стихийное вытеснительное скрещивание происходит при скрещивании местных пчел с завозными породами.
Хочется предостеречь в селекционной работе от необдуманного скрещивания, метизации ценных популяций местных пчел, наилучшим образом приспособленных к условиям среды. Исходя из учета одного лишь показателя — медопродуктивности, /. М. Cornuet (1981) предложил следующий упрощенный план ее повышения.
В каждом поколении отбирают 10 пчелиных семей. От каждой из этих семей получают маток для организации 12 семей-дочерей. Таким образом получают 120 пчелиных семей от 10 основательниц. Эти 120 семей распределяют на четыре точка по 30 семей. По результатам оценки медопродуктивности этих 120 семей отбирают из них 10 для дальнейшей селекции. Отбор основывается как на собственной ценности семьи, так и на средней ценности семей данной группы. Спаривание естественное, но принимаются меры для выращивания большого количества трутней нужного происхождения. Схема данной селекции строится на основе местной популяции с обязательным мечением маток.
J.P.Chaplean (1984), рассматривая различные подходы к селекции пчел, отметил, что их медопродуктивность нельзя повысить, не проводя отбора по качеству потомства. При этом можно подбирать сотни пчелиных семей по длинному списку критериев, и можно ограничиться лишь несколькими признаками и небольшим количеством пчелиных семей (в зависимости от возможностей селекционера). Поскольку контроль зa генотипом трутней осуществить трудно, да и спариваться они могут раз в жизни, приходится, как говорилось выше, отдавать предпочтение селекции по женской линии. Однако и в этом случае можно значительно улучшить качество потомства. Такая селекция дает эффект уже у следующего поколения трутней.
Этот автор предлагает два метода селекции — групповую, когда от одной или нескольких отобранных маток получаются все остальные (и так из поколения в поколение) при естественном спаривании, и групповую с группами. В последнем случае матки в каждой подгруппе — сестры. Ежегодно новая матка-родоначальница выбирается внутри каждой подгруппы и ее дочери продолжают линию. Это, собственно, схемы массового (фенотипического) и генотипического отбора.
J.Vaillqnt (1982) вслед за другими авторами призывает учитывать наследственность трутней и считает, что она имеет не меньшее значение, чем наследственность маток в формировании генотипа женских особей потомства. Это не отрицают и другие селекционеры, однако подобное возможно при наличии надежных изолированных случных пунктов или при использовании инструментального осеменения.
К тому же, занимаясь селекцией трутней, следует иметь в виду, что у них могут быть лишь дочери, деды и внуки. Матки чистой породы производят генетически идентичных трутней той же породы. Если в определенную местность завозят матку другой породы, она дает трутней своей породы, которые будут спариваться с местными неплодными матками, вытесняя прежнюю породу. В одной пчелиной семье самки, происходящие от одного трутня, имеют коэффициент родства 3/8 вместо ]/4 при наличии двух диплоидных родителей, поэтому Лейдлоу предложил называть их суперсестрами. Риск близкородственного скрещивания компенсируется многократным спариванием маток. В противном случае близкородственное скрещивание привело бы к снижению резистентности рабочих пчел к заболеваниям, уменьшению активности их летной деятельности, сокращению продолжительности жизни трутней, появлению генетически «пестрого» расплода. Чтобы этого не случилось, необходимо оперировать несколькими отобранными линиями.
Признаки отбора пчелиных семей для вывода трутней соответствуют таковым при выводе маток.
Пчелиная семья в период максимального развития отстраивает до 14% трутневых ячеек, а в конечном итоге их бывает 2—3% от общего количества рабочих пчел. Если нет надежного изолированного пункта спаривания, необходимо применять следующие приемы: стимуляцию селекционных трутней к вылету в определенное время, когда другие трутни не летают, спаривание вне сезона, создание мощного трутневого барьера за счет преимущественного вывода нужных трутней.
W.G.Rothenbuhler (1980) считает, что любая программа улучшения породы пчел должна состоять из трех этапов: полевых испытаний, селекционной работы и коммерческого производства улучшенных пород пчел. При этом полевые испытания должны проводиться именно в тех районах, где порода будет использоваться. Задача селекционера — спланировать порядок полевых опытов, отобрать пчелиные семьи для селекции, решить, когда использовать инструментальное осеменение, а также как поддерживать и использовать улучшенную породу. По мнению этого автора, успех осуществления программы правильного определения района испытания пчелиных семей зависит от признаков отбора (как правило, основной — медопродуктивность), количества пчелиных семей, испытываемых в каждом поколении (в среднем 50); а также процента из них, необходимого для дальнейшей селекции (автор,советует брать 10%). Также важны организация полевых испытаний оплодотворенных естественным путем маток, продолжение подобной работы в следующем поколении, сравнение новой линии с используемыми на пасеке пчелами, размножение и реализация маток этой линии в случае ее превосходства.
Издавна между селекционерами-пчеловодами идет дискуссия о том, использовать ли чистые породы или межпородные гибриды.
W.С.Roberts, О.Mackensen (1951) показали, что противоречий в этих методах нет. Идеальная система скрещивания — это селекция и поддержание чистых, но неродственных линий и их скрещивание для получения улучшенных гибридов. Выводимые инбредные линии слабее неинбредных, однако их правильные комбинации, как уже говорилось, дают эффект гетерозиса.
Н.Cale, J.Н.Gowen (1956) проанализировали генетический материал из четырех инбредных линий пчел. Ими получены различные результаты, позволяющие сделать вывод о том, что не все гибриды одинаково ценны, и приходится изучать и учитывать комбинационную способность инбредных линий.
При хорошей организации и планировании чистопородного разведения удается получать значительный селекционный эффект без риска метизации и потери ценных породных качеств. Чистопородная селекция основывается на долгосрочных, рассчитанных на несколько лет, комплексных программах.
Селекционные программы, используемые в странах с развитым пчеловодством
Знакомство с программами хотелось бы начать с опыта селекционеров Австрии. Систему племенной работы в этой стране регламентирует Австрийский союз пчеловодов (Н.И.Кривцов, Г.Н.Котова, 1983). Основным принципом его деятельности является тесная связь науки и практики — Института пчеловодства как центра племенной работы и пчеловодов-промышленников на местах. В этом комплексе ежегодно задействовано 1500—2000 пчелиных семей. В Австрии разводят в основном аборигенных краинских пчел, а в северных районах — темных лесных. Племенных маток передают на частные пасеки для испытания. От каждой проверяемой по качеству потомства матки оценивают по 5—15 маток-дочерей. Австрийские селекционеры считают, что лучше сократить число дочерей в группах, но проверить по качеству потомства как можно больше маток-рекордисток. Оценку ведут 1—2 года. Важнейшим селективным признаком при этом является медопродуктивность. Ее учитывают пчеловоды-практики на своих пасеках и по специальной форме передают результаты в институт для анализа. Если медопродуктивность устраивает селекционеров, то приступают к оценке других признаков, и прежде всего чистопородности. Чистопородность по экстерьерным признакам устанавливает институт.
Таким образом, на частных пасеках, расположенных в различных условиях, проводится конкурсное испытание племенного материала, получаемого из института. Данные сравнивают с показателями местных линий. Если в результате оценки на пасеках-испытательницах выявляется ценный для селекции материал, то его передают институту в виде яиц и молодых личинок для последующей репродукции.
Опираясь на сеть пасек-испытательниц, институт ведет 7 линий по 100 пчелиных семей. Спаривание маток институт осуществляет на станциях спаривания, расположенных в изолированных долинах Альп. Рассылаемых маток метят метками различной формы и цвета, обозначающими их происхождение и год рождения. Постоянная пропаганда и реклама результатов селекции способствуют повышенному спросу на племенных маток. Пчеловодам, которые аккуратно и добросовестно присылают сведения о результатах испытания, поставляют маток на льготных условиях.
В ФРГ (Баварский институт пчеловодства) селекционируют линии (штаммы) краинских пчел, которые оценивают на трех испытательных пунктах (К.Weiss, 1982). На каждом пункте испытывают 10 групп по 10 пчелиных семей в каждой. Испытания, не считая год подготовки, когда подсаживают маток, продолжаются 2 года.
Продуктивность оценивают и регистрируют по каждой пчелиной семье отдельно. Детальные отчеты даются за каждый год испытания, а после окончания испытания институт представляет в специальные издания по пчеловодству обобщенный отчет.
В бывшей ГДР селекционеры использовали карнику (G.Pritsch, 1983). Были созданы многочисленные случные пункты с отобранным трутневым составом определенного происхождения. С мест испытания сюда возвращали оцененный племенной материал. Работу с каждой линией обеспечивали 5—6 пунктов, образующих комплекс. Каждый такой комплекс представлял собой замкнутую селекционную систему, что давало возможность исключить бесплановые скрещивания. Всего же действовало 60 случных пунктов, объединенных в 12 замкнутых комплексов. Племенные пчелиные семьи отбирались по принципу чистопородности в соответствии со стандартом. В радиусе 10 км от случного пункта не должно было быть инородного материала
Именно на основе комплексов случных пунктов и строилась работа по планам ступенчатого линейного разведения (линии оценивали в нескольких, не менее чем в трех, поколениях). Здесь же получали необходимое количество чистопородных племенных маток, контролировали кочевки в зонах чистопородного разведения и оценивали чистопородность племенного материала.
В Чехии (V.Vesely, 1982) селекционную работу координирует Институт пчеловодства в Доле. Непосредственное участие в организации этой работы принимает Чешский союз пчеловодов. Все пасеки делятся на следующие категории: областные, признанные, разведенческие, зарегистрированные, товарные.
Областные пасеки относятся к высшей категории. Здесь селекционные достижения внедряются в практику, количество пчелиных семей на таких пасеках должно быть не менее 40.
На признанных пасеках (размером также не менее 40 пчелиных семей) обеспечивается контроль над спариванием маток. Селекцию здесь ведут методом индивидуального отбора. Пчеловоду необходимо доказать свою профессиональную пригодность к работе на признанной пасеке. Это значит, он должен не менее чем в трех поколениях пчел проводить оценку признаков, в том числе и экстерьерных, на соответствие стандарту, обеспечивать контроль над спариванием, а также подтвердить, что в течение двух лет средний показатель медопродуктивности составлял не менее 80% от показателя областной пасеки. Признание его профессиональности действует 10 лет, затем проводят повторную проверку по перечисленным выше критериям.
Разведенческие пасеки размножают племенной материал какой-либо признанной или областной пасеки. Здесь ведут массовый отбор трутней, ежегодно заменяют не менее 60% маток. Такие пасеки также насчитывают не менее 40 пчелиных семей при непременном условии ежегодной продажи не менее 100 маток на сторону. Спаривание маток проводят на территории пасеки. Заявитель на статус разведенческой пасеки представляет карту местности с указанием местонахождения своей пасеки и пасек других пчеловодов в радиусе 5 км, которые будут брать маток именно у него и заменять ими ежегодно не менее 20% маток на своих пасеках.
Зарегистрированные пасеки насчитывают не менее 20 пчелиных семей, на них проводят массовый отбор.
Товарные пасеки могут выращивать маток только для собственных нужд.
В институте и на областных пасеках организована специальная служба, обеспечивающая спаривание привезенных в нуклеусах неплодных маток.
В Румынии (М.Щербан, М.Драган, 1984) по результатам оценки на конкурсах, по данным научных сотрудников, пчеловодов на высокопродуктивных пасеках, ежегодно выявляют маток — рекордисток по медопродуктивности. Оценку пчелиных семей проводят на испытательных и селекционных пасеках, организованных на зональном уровне. Кроме основного признака, медопродуктивности, учитывают яйценоскость, миролюбие, поведение на сотах, ройливость и др. Отличившиеся по комплексу признаков пчелиные семьи передают на элитные пасеки или в центры, предназначенные для создания и совершенствования новых линий или кроссов. Работа на этом уровне относится непосредственно к компетенции института. Элитные пасеки и центры производят материал высокой племенной ценности, они постоянно обмениваются ценными, по собственным результатам и результатам потомства, матками с другими пасеками страны и пасеками-испытательницами.
Линии со стабильными признаками и лучшие кроссы представляют специальной комиссии для официального признания и включения в книгу республиканского центра селекции.
Представление о системе селекционной работы в США можно получить, познакомившись с работой фирмы «Дадан и сыновья» и генетико-селекционной лаборатории в Батон-Руже (П.Уйтрелл, 1976; А.Харнаж, Т.Росс, Т.Дадан, Г.К.Кейл, 1976; G.Claerr, 1977).
Программа селекции в США предусматривает получение и сохранение инбредных линий и широкое использование в производстве межлинейных кроссов. Для получения 2-, 3-, 4-линейных кроссов используют маток с высокой степенью родства.
Получение планомерных гибридов стало возможным в США с развитием техники инструментального осеменения пчелиных маток (F.Е.Moeller, 1976). Для того чтобы матки также были гибридами, проводили многократные (3—6) межпородные скрещивания. Инбредные линии были произведены от нескольких пород, затем их комбинации испытывались на продуктивность. Всей программе селекции сопутствовал непрерывный отбор по медопродуктивности, устойчивости к американскому гнильцу и некоторым другим характеристикам (злобливость, ройливость, склонность к прополисованию, качество расплода, характер печатки меда, окраска тела).
Чтобы сохранять преимущества гибридов, необходимо поддерживать в чистоте инбредные линии и всякий раз получать гибрид первого поколения. Поддерживать же инбредные линии, имеющие низкую жизнеспособность, непростая задача. Поэтому следует избегать высокой степени инбридинга исходных пород.
Маток, дающих пчел межлинейных кроссов, распределили между участвующими в программе пчеловодами-селекционерами, которые их испытали и отобрали племенной материал для последующих межлинейных скрещиваний. Чистота кроссов гарантировалась инструментальным осеменением.
Для выполнения комплексных селекционных программ считается целесообразным иметь специальную пасеку с лабораторией сохранения и поддержания инбредных линий, пасеку для получения межлинейных кроссов первого поколения и сеть пасек для оценки их селекционной ценности. Так, широко распространенный в США гибрид Старлайн основывается на четырех инбредных линиях итальянских пчел. Ежегодно выводят новые линии, которые сравнивают с уже существующими. Оказавшиеся лучшими в результате оценки линии становятся источником получения маток и трутней для очередного поколения. Этот племенной материал пчеловоды-селекционеры возвращают фирме для получения коммерческих кроссов. Такая кооперация фирмы с пчеловодами-селекционерами очень эффективна.
Обобщая зарубежный опыт, следует отметить рациональные стороны чистопородной селекции.
Работу репродукторов, как правило, курируют научные учреждения селекционного профиля, что позволяет размножать племенной материал, постоянно улучшаемый в процессе селекции.
Существует постоянная взаимосвязь между центрами селекции и репродукторами, с одной стороны, и заказчиками маток — с другой. Это дает возможность анализировать результаты, получаемые на местах, и, таким образом, отбирать наиболее ценный материал для дальнейшей селекции и репродукции. Благодаря организации обратной связи небольшой коллектив селекционеров может решать задачи крупномасштабной селекции.
Селекционная работа проводится на основе относительно крупных массивов пчел одного происхождения, что исключает возможность бесплановой метизации. В репродукторах чистота племенного материала поддерживается периодическим инструментальным осеменением, если нет надежных изолированных случных пунктов.
В основе селекции — четкие программы, регламентированные соответствующими органами.
Методы селекционной работы в России
В нашей стране чистопородная селекция строится либо на основе массового, фенотипического отбора, либо с использованием индивидуального, генотипического отбора с проверкой маток по качеству потомства и выведением линий.
Массовый отбор является наиболее доступным и испытанным способом улучшения хозяйственных и племенных качеств пчелиных семей. Его сущность заключается в планомерной выбраковке худших пчелиных семей и интенсивном размножении лучших. Как уже говорилось, отбор ведется по медопродуктивности, однако учитываются и такие хозяйственно полезные признаки, как зимостойкость, устойчивость к заболеваниям, плодовитость маток и сила пчелиных семей, ройливость, воскопродуктивность и т. д.
В конце сезона все пчелиные семьи оценивают по перечисленным признакам и распределяют на 3 группы. В первую группу выделяют 10—15% лучших пчелиных семей, в 1,5—2 раза превосходящих средние показатели пасеки по медопродуктивности, хорошо перезимовавших, без признаков заболеваний, с высоколодовитыми матками и т. д. Кроме того, семьи пчел, выделяемые в первую (племенную) группу, должны быть чистопородными, т. е. отвечать по всем своим признакам типу дайной породы. Так, у среднерусских пчел окраска тела не должна иметь желтизны, печатка меда должна быть белой («сухой») и т д.
Ко второй группе, самой многочисленной, относят пчелиные семьи со средними показателями. К третьей группе (10—15% от общего числа пчелиных семей) относят худших по остальным показателям и наименее продуктивных. Семьи третьей группы необходимо выбраковывать сразу же, т. е. в конце сезона, или в крайнем случае в следующий сезон.
В следующий сезон в качестве племенных используют семьи первой группы. В семьях второй группы сменяют маток на маток, выведенных от семей первой группы. Для смены старых маток в семьях первой группы и для подсадки во вновь формируемые отводки используют также только маток, выведенных от лучших семей первой группы.
В выделенных из первой группы отцовских семьях создают наилучшие условия для выращивания трутней— подставляют 1—2 трутневых сота, тщательно утепляют гнезда, систематически дают стимулирующую подкормку. Одновременно с этим применяют все меры, чтобы не допустить вывода трутней во всех остальных семьях пасеки.
Улучшение породы будет более успешным, если такую же племенную работу ведут и на окружающих пасеках, расположенных в зоне с радиусом до 12—15 км. В противном случае возможно нежелательное спаривание значительной части маток с трутнями из малопродуктивных семей соседних пасек.
Целесообразно выводить молодых маток от пчелиных семей, неродственных отцовским, а также через 1 - 4 года производить обмен племенными пчелиными семьями между пасеками, разводящими одну и ту же породу пчел и находящимися друг от друга не ближе, чем в 25—30 км, чтобы избежать родственного спаривания маток и трутней.
Массовый отбор, проводящийся систематически из поколения в поколение, улучшает также племенные качества пчелиных семей, т. е способность устойчиво передавать своим дочерним семьям высокую продуктивность и другие ценные хозяйственно полезные признаки.
Описанная схема применима, разумеется, к пчелам всех пород. Неоднократно отмечалось, что массовый отбор на пасеках среднерусских пчел приводил к значительному повышению их медопродуктивности. Так, Н.Солодкова (1951), Т.С.Жданова (1970) сообщали, что в результате массового отбора среднерусские пчелы татарской популяции при благоприятных условиях климата и взятка могут собирать за сезон до 100 кг меда на каждую пчелиную семью.
Индивидуальный генотипический отбор — более высокая форма племенной работы. Однако исходные позиции остаются такими же, как и при применении массового отбора. Индивидуальную селекцию ведут, как правило, в научно-исследовательских учреждениях по пчеловодству, располагающих необходимой для этого базой и силами.
Имеются примеры чистопородной селекционной работы со среднерусскими пчелами. Так, Н.Н.Гранкин (1978), занимающийся селекцией среднерусских пчел орловской местной популяции, выделил линии № 39 и 7, превосходящие контроль по медовой продуктивности соответственно на 28,5 и 39,4%, а по максимальной яйценоскости — на 39,4 и 32,5%. Этим селекционером апробированы и некоторые варианты внутрипородных межлинейных гибридов среднерусских пчел, также оказавшихся эффективными.
И.В.Шафиков (1978) проводил похожую работу с аборигенной бурзянской бортевой пчелой. В результате селекций им была выделена линия № 79 с повышенной на 25—35% плодовитостью маток, медовой и восковой продуктивностью пчелиных семей. В абсолютном выражении в проведенных испытаниях пчелиные семьи с матками-дочерьми, родоначальницы линии № 79, собрали меда на 8,3 кг больше, чем семьи контрольной группы. Пчелиные семьи с матками-внучками линии № 79 превышали семьи контрольной группы на 9 кг.
В отличие от других отраслей животноводства, где работа ведется, как правило, с мужскими линиями, в пчеловодстве основное внимание уделяется маткам и, соответственно, женским линиям. Это вызвано такими биологическими особенностями медоносных пчел, как большая плодовитость маток (до 2000 и более яиц в сутки), их скороспелость (от племенной матки за летний сезон можно получить и дочерей и внучек). Принимается во внимание и то, что трутни способны к спариванию лишь один раз.
Начало работы по проверке качества потомства пчелиных маток было положено исследованиями пчело-генетической лаборатории сектора генетики и селекции Всесоюзного института животноводства (ВИЖ) в 1930—1935 гг. под руководством академика А.С.Серебровского. После значительного перерыва селекционные работы были возобновлены, и в 70-е гг. был успешно применен метод проверки маток по качеству потомства при селекции дальневосточных, серых горных кавказских, карпатских пчел (Г.А.Аветисян, В.А.Губин, И.К.Давыденко, 1969; Ю.И.Макаров, 1969; В П.Кидалов, 1971, и др.). Все селекционеры отмечали большие возможности линейного разведения пчел различных пород с оценкой маток по качеству потомства, однако исследователи в большинстве случаев ограничиваются получением и испытанием одного-двух поколений.
СЕЛЕКЦИЯ СРЕДНЕРУССКИХ ПЧЕЛ
Как уже было отмечено, исходным материалом для селекции пчел среднерусской породы послужили 92 пчелиные семьи татарской популяции, завезенные из Сабинского района Татарии. Кстати, эта популяция в настоящее время успешно разводится в Мамадышском и Сабинском районах Татарии, где организован заказник среднерусских пчел.
В селекционном процессе участвовали только чистопородные пчелиные семьи (табл. 10). Племенное ядро линии № 44 было размещено в условиях относительной пространственной изоляции (радиусом около 3 км), и поэтому для сохранения чистопородности использовались дополнительно такие приемы, как изоляция во времени, создание мощного трутневого барьера, жесткий отбор по окраске хитина и экстерьерным признакам пчел. В результате этого был сформирован относительно устойчивый чистопородный массив среднерусских пчел в районе опытного поля.
 Ниже приводятся значения наиболее характерных экстерьерных признаков пчел исходной популяции по сравнению с усредненными данными восьмого и девятого поколений линии № 44 (осенняя популяция), полученными на 10 производственных пасеках массива (табл. 11).
Ниже приводятся значения наиболее характерных экстерьерных признаков пчел исходной популяции по сравнению с усредненными данными восьмого и девятого поколений линии № 44 (осенняя популяция), полученными на 10 производственных пасеках массива (табл. 11).
Как видно из табл. 11, за 19 лет селекционной работы изменения экстерьерных признаков не произошло, они остались в рамках стандарта породы. Отмечено некоторое увеличение их фенотипической изменчивости (по значениям Cv%), однако значительную долю этого увеличения можно отнести за счет издержек при расчетах. Многолетний опыт свидетельствует о том, что даже в условиях недостаточной пространственной изоляции и при большой насыщенности пчелиных семей можно успешно сохранять и размножать чистопородный материал.
Основываясь на сделанных ранее расчетах, основным селективным признаком выбрали медопродуктивность. Поскольку зимостойкость связана с медопродуктивностью положительной корреляцией, она достаточно высока у среднерусских пчел, и задача состоит лишь в контроле за ее уровнем, а не в селекционном улучшении этого признака. К тому же отбор на медопродуктивность косвенно способствует отбору на зимостойкость.
Яйценоскость пчелиных маток (как среднесуточная, так и максимальная) также связана с медопродуктивностью положительной корреляцией (г=0,39 — 0,91), но этот признак может считаться, в зависимости от цели, и самостоятельным селективным (например, в хозяйствах разведенческого направления).
На первых этапах селекции отбор на медопродуктивность был очень жестким: на племя оставляли пчелиные семьи, превосходящие показатели сверстниц на 2q. В дальнейшем, по мере того как исчерпывалось генотипическое разнообразие, отбирали просто лучшие по медопродуктивности пчелиные семьи. В каждом поколении для репродукции последующего поколения отбирали 3—5 маток-рекордисток.
Селекционная работа проводилась на фоне относительно скудного главного медосбора с гречихи и сорно-полевого разнотравья, свойственного для центральных областей Нечерноземья, однако схема опыта дала возможность сделать объективные выводы об относительной ценности испытываемых по качеству потомства пчелиных семей аутбредной по трутням линии № 44.
В табл. 12 приведены данные о медопродуктивности племенных пчелиных семей этой линии.
 Для репродукции очередного поколения линии, за исключением первых двух, отбирали 3—5 пчелиных семей. Показатели медопродуктивности различны по годам, так как сильно зависят от медосборных условий сезона.
Для репродукции очередного поколения линии, за исключением первых двух, отбирали 3—5 пчелиных семей. Показатели медопродуктивности различны по годам, так как сильно зависят от медосборных условий сезона.
При планировании опыта по Е.К.Меркурьевой (1970) определили численность дочернего потомства для оценки маток по медопродуктивности их дочерей. При q признака равной 5 кг и точности А в размере 3 кг, при критерии достоверности t 0,95=1,96 она равна 21 пчелиной семье:


 В случае применения индивидуального отбора пчелиных семей для получения трутней и гомогенного подбора по яйценоскости результаты, очевидно, были бы еще значительнее, хотя и при использованных селекциониых методах они говорят сами за себя.
В случае применения индивидуального отбора пчелиных семей для получения трутней и гомогенного подбора по яйценоскости результаты, очевидно, были бы еще значительнее, хотя и при использованных селекциониых методах они говорят сами за себя.
Таким образом, яйценоскость пчелиных маток, является важным селективным признаком, имеющим как самостоятельную селекционную значимость, так и пригодным для проведения косвенного отбора на медопродуктивность. Уже при жестком отборе можно довести уровень яйценоскости до предельных значений, превышающих стандарт породы.
 В табл. 18 обобщен селекционный эффект, полученный по всем селективным признакам. Данные показывают, что в среднем за 9 поколений селекции превышение над уровнем исходной популяции составило по медопродуктивности 57,9%, по максимальной яйценоскости 40,2% и по среднесуточной — 64,4%. Превосходство над уровнем селективных признаков по сравнению с матками-сверстницами несколько ниже (28,3—41,9%), причем к F-7 оно становится незначительным (4,2—5,0%). Очевидно, до ничтожно малых значений снизился уровень генотипического разнообразия признаков. Это же подтверждает и анализ степени наследуемости селективных признаков. Их значения с 8— 26% в F-1 достигли 1—3% в F-7. Значительно снизились и значения коэффициентов фенотипической и генотипической изменчивости.
В табл. 18 обобщен селекционный эффект, полученный по всем селективным признакам. Данные показывают, что в среднем за 9 поколений селекции превышение над уровнем исходной популяции составило по медопродуктивности 57,9%, по максимальной яйценоскости 40,2% и по среднесуточной — 64,4%. Превосходство над уровнем селективных признаков по сравнению с матками-сверстницами несколько ниже (28,3—41,9%), причем к F-7 оно становится незначительным (4,2—5,0%). Очевидно, до ничтожно малых значений снизился уровень генотипического разнообразия признаков. Это же подтверждает и анализ степени наследуемости селективных признаков. Их значения с 8— 26% в F-1 достигли 1—3% в F-7. Значительно снизились и значения коэффициентов фенотипической и генотипической изменчивости.
Из этих данных можно сделать вывод о том, что такие признаки, как медопродуктивность и особенно яйценоскость среднерусских маток, отзывчивы на селекцию. Наибольший эффект отмечен в F-1—F-4. Очевидно,в селекции среднерусских пчел целесообразно иметь дело с короткими линиями (до F-1—F-4).
 Одним из аспектов селекционной работы может быть селекция линий среднерусских пчел, специализированных на опылении отдельных энтомофильных культур, в частности гречихи.
Одним из аспектов селекционной работы может быть селекция линий среднерусских пчел, специализированных на опылении отдельных энтомофильных культур, в частности гречихи.
Флоромиграцию пчелиных определяют цвет, запах цветков, наличие нектара и пыльцы и их доступность. Поскольку нектар растений различных семейств отличается по составу сахаров, а также по аминокислотному составу, то эти факторы также могут влиять на флоромиграцию и играть определенную роль в коэволюции растений и опылителей (Харборн, 1985).
У одиночных пчел наблюдается довольно узкая приспособленность к растениям определенных семейств. Так, представители рода Andrene посещают в основном орхидные.
Общественные пчелы, и в частности Apis mellifera, посещают довольно широкий спектр энтомофильных растений, отдавая предпочтение семействам губоцветных, норичниковых и бобовых, цветки которых имеют главным образом голубой и желтый цвета. Однако и внутри вида медоносной пчелы отмечается избирательность различных пород (рас) пчел по отношению к тем или иным растениям. Еще И.Н.Клинген (1912) убедительно показал преимущество длиннохоботных серых горных кавказских пчел в опылении красного клевера.
Ш.О.Гасанов (1970), подтверждая данные предыдущих исследований о наличии флороспециализации у пчел разных пород (рас), показал, что среднерусские пчелы предпочитают гречиху и липу, серые горные кавказские— клевера, сурепку, фацелию.
Принимая наличие флороспециализации как бесспорный факт, селекционеры стали предпринимать попытки выведения линий пчел, специализированных на опылении отдельных культур: люцерны (Макензен и Най, 1967; Cale, 1971; И.К.Давыденко и др., 1980), красного и белого клеверов (Холм, 1971).
Совместно с Д.В.Гирником мы изучили возможность селекции пчел на опыление гречихи. Наши исследования косвенно подтверждают вывод: пчелы, отловленные на цветках гречихи, по своим экстерьерным признакам ближе к стандарту среднерусской породы, чем пчелы, посещающие цветки других растений (табл. 19).
Как правило, при благоприятной погоде пчелы хорошо посещают гречиху. Однако бывают сезоны, когда даже при хорошей вегетативной массе цветки плохо выделяют нектар, что сказывается отрицательно на ее урожайности.
Возникает необходимость усиления опылительной деятельности пчел на этой культуре.
Уже подчеркивалось, что выбор главного селективного признака имеет определяющее значение. Известно, что с одних растений пчелы той или иной породы предпочитают сбор нектара, с других — сбор пыльцы, с третьих — одинаково хорошо собирают и пыльцу и нектар одновременно. Если пчелы испытываемой породы охотно собирают пыльцу с данной культуры, то в качестве основного селективного признака следует брать именно его. Если же они охотно собирают и пыльцу и нектар, то тоже лучше брать в качестве основного селективного признака склонность собирать пыльцу, так как провести ботанический анализ пыльцы проще, чем сделать подобный анализ нектара.
Мы проследили за работой на цветках гречихи пчел, собирающих пыльцу, и пчел, собирающих нектар, а также учли завязываемость семян гречихи после опыления их пыльце- и нектаро сборщицами. Эффективность опыления цветков гречихи пчелами разной специализации составила (%):
Пыльценектаросборщицы (54 пчелы) ........... 78,3 (94 завязавшихся зерна)
Нектаросборщицы (29 пчел) ............... .21,7 (26 завязавшихся зерен)
Всего (83 пчелы) .................... 100 (120 завязавшихся зерен)
Около 80% опылительной работы на цветках гречихи приходится на пчел, собирающих одновременно и пыльцу и нектар, т. е. на пыльце-нектаро сборщиц. Полученные данные указывают на то, что в качестве главного селективного признака при выведении специализированных («гречишных») линий, следует использовать склонность отдельных семей пчел среднерусской породы собирать пыльцу этой культуры.
 Опыт показал, что степень склонности к сбору гречишной пыльцы наследуется матками-дочерьми. Дочерние семьи с матками, полученными от родителей с высокой степенью проявления этого признака, собирали достоверно больше гречишной пыльцы (49,34=5,23), чем пчелиные семьи, полученные от родителей с низкой склонностью собирать пыльцу с гречихи (26,1 ±10,5%).
Опыт показал, что степень склонности к сбору гречишной пыльцы наследуется матками-дочерьми. Дочерние семьи с матками, полученными от родителей с высокой степенью проявления этого признака, собирали достоверно больше гречишной пыльцы (49,34=5,23), чем пчелиные семьи, полученные от родителей с низкой склонностью собирать пыльцу с гречихи (26,1 ±10,5%).
Мы провели индивидуальный отбор пчелиных семей из группы с высокой склонностью к сбору гречишной пыльцы и из группы с низкой склонностью и вывели от них второе поколение маток. Индивидуальный отбор пчелиных семей для вывода трутней по данному селективному признаку не проводили, и спаривание было свободным. И в этом случае имело место наследование склонности к сбору гречишной пыльцы. Однако разница между указанными группами была значительно меньше и составляла лишь 7%. Полученные данные говорят о том, что при имеющемся уровне наследуемости (/г2=0,25) массовый фенотипический отбор на «гречишность» имеет определенный смысл, но для получения большего эффекта необходимо применять генотипический отбор и подбор, что возможно только при условии строгого контроля над спариванием..
Таким образом, было подтверждено наличие у среднерусских пчел флороспециализации к гречихе и показана принципиальная возможность селекции линий, специализированных на опылении этой культуры.
Принципиальная схема чистопородной селекции
На основании изученной литературы и проведенных исследований можно предложить следующую примерную схему чистопородной селекции с породами пчел.
Ознакомление с генофондом и ресурсами породы, выявление в результате обработки литературы, анкетирования, а также экспедиционных поисков сохранившихся в чистоте в пределах ареала естественного обитания чистопородных пчел (популяции, экотипы, отдельные пасеки). Уточнение по комплексу экстерьерных, хозяйственно полезных, биологических признаков их породной принадлежности. Изучение имеющейся у пчеловода документации относительно происхождения и продуктивности пчелиных семей. Завоз маток (желательно одновозрастных) или пчелопакетов, пчелиных семей из выявленных пунктов в научно-исследовательское или опытное учреждение по пчеловодству. Организация на месте достаточно больших групп пчелиных семей нужного происхождения с использованием инструментального осеменения или изолированных случных пунктов (3—4 года).
Сравнительное испытание завезенных из разных мест популяционных групп пчелиных семей по комплексу хозяйственно полезных и биологических признаков. Выделение для дальнейшей работы перспективных популяций, обладающих комплексом ценных хозяйственно полезных признаков и пригодных для закладки на их основе заводских линий (представляют интерес и популяции, характеризующиеся каким-либо одним ценным качеством для закладки специализированной линии). Установление делового контакта с учреждениями и пчеловодами-любителями тех мест, из которых были завезены пчелы, оказавшиеся лучшими в сравнительном испытании, с целью осуществления повторных завозов для «освежения крови». Проведение необходимых мероприятий, препятствующих метизации аборигенных пчел в этих местах (создание заказников, заповедных зон и т. д ) (2—3 года).
Размножение пчелиных семей лучших популяций (желательно в условиях пространственной изоляции), организация популяционных пасек. Осуществление контроля над спариванием известными методами (изоляция в пространстве и времени, применение инструментального осеменения, двукратная смена маток). Репродуцирование наиболее чистопородного материала в центре популяционного массива, где также осуществляется репродукция маток как для расширения массива, так и для реализации на сторону.
Проведение в популяционных массивах постоянной работы по селекционному улучшению и контролю за чистопородностью, фенотипическая оценка пчелиных семей популяционных пасек, отбор предполагаемых родоначальниц линий и получение от них групп маток-дочерей для проверки их по качеству потомства. Анализ фенотипической и генотипической изменчивости, наследуемости, повторяемости, предварительный прогноз эффекта селекции. Изучение комплекса корреляционных связей между хозяйственно полезными и биологическими признаками с целью применения косвенного отбора, играющего особо важную роль в селекционном улучшении признаков с низкой наследуемостью.
Выведение аутбредных (по типу заводских) линий пчел, характеризующихся четко выраженным развитием основных селективных признаков (медопродуктивности, яйценоскости) при сохранении других хозяйственно полезных признаков на уровне стандарта для породы. Продолжение линий в 3—4 поколениях, анализ генетико-популяционных параметров, выяснение целесообразности получения последующих поколений и закладки новых линий.
Получение товарных внутрипородных гибридов (межпопуляционных и межлинейных) с целью хозяйственного использования эффекта микрогетерозиса. Испытание полученных гибридов на сочетаемость с целью последующего использования лучших вариантов в промышленных внутрипородных скрещиваниях.
Фактический объем внедрения плодных маток линии № 44 составил 10 тыс. шт., и следовательно, экономический эффект, выраженный через прибавку медопродуктивности, равен 3599 тыс. руб. По нашим расчетам, с расширением объема репродукции до удовлетворения полного спроса эффект может составить около 26 млн руб. (в ценах 1990 г.).
ИСПОЛЬЗОВАНИЕ СРЕДНЕРУССКИХ ПЧЕЛ В МЕЖПОРОДНЫХ ПОМЕСЯХ
В настоящее время можно считать бесспорно доказанным факт проявления гетерозиса при скрещивании пчел некоторых завозных пород с местной среднерусской (А.Н.Брюханенко, 1929; А.А.Курочкин, 1030; М.3.Краснопеев по В.В.Алпатову, 1948 Г.Д.Билаш, 1956; Г.Ф.Таранов, 1956; А.Н.Биктимиров, 1963; А.Я.Шекшуев, 1967; А.В.Бородачев, В.Т.Бородачева, 1982, и др.).
Перечисленные авторы, как правило, использовали маток завозной породы, а трутней — местной среднерусской. Полученные помеси наследовали такие качества исходных пород, как хорошая зимостойкость, кормодобывающая активность, работоспособность. Эффект гетерозиса проявлялся на качестве рабочих пчел. Вместе с тем при простых скрещиваниях генотип матки не затрагивается, и, следовательно, эффект гетерозиса по яйценоскости маток, а также улучшающее действие другой породы не проявляются.
В то же время матки среднерусской породы обладают высоким потенциалом яйценоскости, который можно было бы использовать в результате скрещивания для усиления этого признака у пород пчел с низкой яйценоскостью.
Помесными матками будут дочери маток от простого скрещивания, спарившиеся с трутнями другой породы, если же их спарить с трутнями третьей породы, рабочие пчелы от них будут сложными трехкратными помесями. Впрочем, помесными будут любые матки-дочери простых помесей. Но если не ставится цель использования в генотипе рабочих пчел третьей породы, можно провести возвратное скрещивание с трутнями одной из уже использованных пород, что повлечет за собой увеличение доли «кровности» этой породы в генотипе пчел до 75%.
Подобные сложные (трехкратные) межпородные помеси с участием среднерусской породы были испытаны в 1979—1983 гг. в Краснодарском крае (Майкопский и Краснополянский опорные пункты пчеловодства) и Башкирии (Башкирская опытная станция пчеловодства). Кроме среднерусской породы (Ср) в помесях использовали пчел кавказской (Кав), итальянской (Ит), карпатской (Кар), краинской (Кр) пород и дальневосточных пчел (Дв). Пчелы среднерусской породы являлись обязательным компонентом сложных помесей. В качестве контрольных были использованы местные породы пчел (серая горная кавказская на юге и среднерусская в Башкирии).
Контроль над спариванием при получении сложных помесей осуществляли или методом инструментального осеменения в НИИ пчеловодства, или в результате использования мощного трутневого барьера в зонах чистопородного разведения на местах.
Среди прочих задач было и выяснение влияния высокояйценоских пород пчел (в частности, среднерусской) на генотип поместных маток с целью увеличения их яйценоскости и, соответственно, медопродуктивности.
В табл. 20 приведены данные о среднесуточной яйценоскости маток перед началом главного медосбора.
 На Краснополянском опорном пункте
На Краснополянском опорном пункте
пчеловодства (ОПП) отмечено очевидное превосходство помесных маток над серыми горными кавказскими, наиболее высокая яйценоскость зафиксирована в варианте ИтХ ХСрХКав за счет высокой яйценоскости двух пород (итальянской и среднерусской), определяющих генотип маток.
На Башкирской ОСП в лесостепной зоне наблюдалось некоторое превосходство (12,2% в среднем за два года) помесных маток варианта ИтХКарХСр над среднерусскими за счет положительного влияния высокояйценоской итальянской породы, а в условиях степи в 1983 г. помесные матки без участия в их генотипе среднерусской породы даже не смогли сравняться в яйценоскости с чистопородными среднерусскими.
На Майкопском ОПП было выявлено убедительное превосходство яйценоскости помесных маток с участием среднерусской и итальянской пород над чистопородными серыми горными кавказскими, хотя доля «кровности» среднерусской породы в генотипе маток в 1980 г. составляла всего лишь 25%.
В табл. 21 показана сила пчелиных семей с матками-помесями на медосборе. Здесь также хорошо прослеживается положительное влияние наследственности среднерусской и итальянской высокояйценоских пород.


Что же касается Майкопского ОПП, то наиболее продуктивными оказались помеси с долей «кровности» среднерусской породы в генотипе рабочих пчел 50%; в 1982 г. эти пчелиные семьи собрали в среднем по 64,2 кг меда, что для местных условий можно считать значительным достижением.
Изучение характера корреляций между такими признаками помесей, как сила пчелиных семей на начало главного медосбора и их медопродуктивность, представляет интерес. Мы изучали этот вопрос в процессе испытания различных вариантов межпородного скрещивания с участием среднерусских пчел, проводимого в различных природно-климатических зонах.
Полученные данные (табл. 23) показывают, что характер корреляций силы семей и медопродуктивности различен в зависимости от происхождения пчелиных семей, места и года испытаний.

Зав. редакцией А.Н.Ивлев Художник Н.В.Медведева Художественный редактор В.В.Быков Технический редактор Л.П.Никитина Корректор В.В.Безымянская Слайды на обложке И.Н.Линича
В книге доктора сельскохозяйственных наук Н. И. Кривцова приведен богатейший материал селекционной работы с пчелами среднерусской породы, а также убедительно продемонстрировано преимущество этой породы для разведения в центральных и северных районах России.
Рассчитана как на ученых-селекционеров, так и на широкий круг пчеловодов-любителей.
 СОДЕРЖАНИЕ
СОДЕРЖАНИЕПредисловие от редакции
Аборигенные пчелы России
Особенности среднерусских пчел
Популяции среднерусских пчел — исходный материал для селекции
Экстерьерные признаки среднерусских пчел
Хозяйственно полезные признаки пчелиных семей различных популяций
Теоретические основы селекции среднерусских пчел
Селектируемые признаки: наследуемость,изменчивость,повторяемость
Фенотипическая корреляция
Организация, методы и программы селекции пчел
Методы селекционной работы
Селекционные программы, используемые в странах с развитым пчеловодством
Методы селекционной работы в России
Селекция среднерусских пчел
Принципиальная схема чистопородной селекции
Использование среднерусских пчел в межпородных помесях
ЗаключениеРедактор В.В.Александрова
© Н.И.Кривцов, 1995
Предисловие от редакции
В основу этой книги положены многолетние научные исследования, проведенные НИИ пчеловодства (а ранее Всесоюзным НИИ пчеловодства), с целью изучения пчел среднерусской породы и их популяций.
Автор — директор НИИ пчеловодства, доктор сельскохозяйственных наук Н.И.Кривцов — приводит богатейший материал селекционной работы с пчелами среднерусской породы, а также убедительно демонстрирует ее преимущества. Особенно перспективно разведение среднерусских пчел в центральных и северных регионах России, где предпочтение отдавалось завозным южным породам пчел, которые плохо приспособлены для климатических и других условий этих территорий.
Автор искренне благодарит своих коллег — Г.С.Балакиреву, А.В.Седых, T.Н.Оринича, В.Н.Власова, И.Я.Джулая, В.М.Виноградову и М.Д.Большакову за помощь в работе и надеется, что книга будет полезна как ученым-селекционерам, так и пчеловодам-любителям.
АБОРИГЕННЫЕ ПЧЕЛЫ РОССИИ
Особенности среднерусских пчел
Пчелы среднерусской породы (среднеевропейские, темные лесные, собственно медоносные — Apis mellifera mellifera) заселяли отросшие в послеледниковое время леса, адаптировались к природно-климатическим условиям и ко времени появления в западной и центральной Европе человека были аборигенами лесистых равнин. Первоначально пчелы распространились севернее и западнее Альп (Швейцария, Франция, Германия, Центральная Россия). Естественное расселение среднерусских пчел дошло, по-видимому, до Урала.
В дальнейшем, по мере развития пчеловодства как важной сельскохозяйственной отрасли, пчелы среднерусской породы были завезены в Сибирь, на Алтай, в Забайкалье, а также в Северную и Южную Америку.
----------------------
По свидетельству Е.М.Баранцевича (1898), одно только бортевое пчеловодство в XVIII веке в Европейской России «за покрытием всех домашних расходов в меде и воске» давало доход 1 млрд руб. ассигнациями. Развитию пчеловодства в значительной мере способствовали незначительная плотность населения, богатейшая растительность. Начиная с 50° с. ш. и южнее — всюду на Руси занимались пчеловодством.
----------------------
По свидетельству Е.М.Баранцевича (1898), одно только бортевое пчеловодство в XVIII веке в Европейской России «за покрытием всех домашних расходов в меде и воске» давало доход 1 млрд руб. ассигнациями. Развитию пчеловодства в значительной мере способствовали незначительная плотность населения, богатейшая растительность. Начиная с 50° с. ш. и южнее — всюду на Руси занимались пчеловодством.
Еще Н.М.Витвицкий (1835), рассуждая об особенностях пчел различного происхождения, писал: «Лучшую же породу их составляют боровки, или боровые, дикие, скрывающиеся от людей в наших лесах и работающие для себя в дуплах гнилых деревьев». Он отмечал способность лесных пчел переносить суровые зимы и пути увеличения продуктивности и доходности пчеловодства видел в обновлении пчельников лесными роями. Лесные пчелы, по словам автора, «всегда резвы, трудолюбивы и прочны».— Примеч. авт.
Большие лесные массивы благоприятствовали пчеловодству. Древнегреческий историк Геродот отмечал, что земли наших предков когда-то были совершенно непроходимы из-за обилия в лесах пчел.
Побывавший в русских лесах Иовий Новокамский писал, что «самая верная жатва получается от воску и меду, ибо вся страна преисполнена плодовитыми пчелами. По лесам и весьма густым рощам здесь можно неоднократно видеть превосходные рои висящих на деревьях пчел, здесь часто находят огромные количества сотов, скрытых в деревах, а в удивительной толщины древесных пнях находят иногда превеликие озера меду» (по А.Морозову, 1978).
Н.М.Витвицкий утверждал, что бортничество и бортевое пчеловодство были некогда для нашего народа тем, чем сейчас является хлебопашество. «Мед и воск были золотом прадедов и дедов наших»,— писал он. В период расцвета бортевое пчеловодство, базировавшееся преимущественно (если не полностью) на среднерусских пчелах, насчитывало в своих угодьях сотни миллионов пчелиных семей, дававших ежегодно почти по 9 млн пудов воска (147,42 млн. кг) и по 600 млн пудов меда (982,8 млн. кг).
Среднерусские пчелы не потеряли своего значения и в настоящее время. Эта порода была рекомендована для разведения в 63 регионах страны, причем в 19 из них, преимущественно северных, она оказалась единственно возможной. Эволюция среднерусских пчел проходила в суровых природно-климатических условиях, в результате чего в процессе приспособления к этим условиям у них появились определенные особенности, закрепившиеся в процессе естественного отбора и отличающие их от пчел других пород. Одним из характерных признаков среднерусских пчел следует считать их поведение при осмотре гнезда. Среднерусские пчелы или «сплывают» к нижнему бруску рамки и повисают гроздьями, или ведут себя слишком возбужденно, быстро перемещаются по соту, взлетают, что очень затрудняет работу с ними. Наряду с этим среднерусские пчелы отличаются сильной злобливостью, и это также является их отрицательным качеством. В периоды отсутствия медосбора даже такие средства защиты от ужалений, как халат, лицевая сетка, обработка дымом, часто оказываются неэффективными.
Большую агрессивность среднерусских пчел по сравнению с кавказскими отмечал, в частности, последовательный сторонник их разведения профессор Г.А.Кожевников (1929). Но он считал, что не все среднерусские пчелы одинаковы в этом отношении.
Ю.В.Страйгис (1974) и Е.М.Петров (1983) указывали на то, что среди местных среднерусских пчел Литвы и Башкирии выделяются семьи повышенной злобливости, которые вместе с тем лучше поддерживают чистоту в улье, более активны и продуктивны.
К сожалению, до сих пор не было предпринято попыток селекции среднерусских пчел на незлобливость, хотя Ф.Руттнер (1968) считал, что такая работа может быть более эффективной, чем отбор по сложным хозяйственно полезным признакам.
Пчелы среднерусской породы печатают мед светлой, «сухой» печаткой, оставляя между восковой крышечкой и поверхностью меда пространство, заполненное воздухом. Это также важный признак породы. У серых горных кавказских и желтых кавказских пчел печатка меда темная, «мокрая».
Для семей среднерусских пчел характерна также нетерпимость маток по отношению друг к другу, в то время как в семьях южных пород нередко наблюдается длительное сожительство в улье двух маток.
Среднерусские пчелы в отличие от пчел южных пород к воровству мало склонны и хуже защищают свое гнездо от пчел-воровок. Возможно, что меньшая склонность к воровству объясняется их слабой предприимчивостью в отыскании источников медосбора. Слабую склонность к воровству следует считать положительным качеством, так как воровитые пчелы затрудняют работу пчеловода на пасеке, а во время подкормок полностью «ограбляют» слабые семьи.
Многие исследователи отмечают у среднерусских пчел высокую склонность к роению. В отдельные годы на пасеках среднерусских пчел до 50—100% пчелиных семей приходят в роевое состояние, а при затянувшемся слабом поддерживающем медосборе начинается бурное роение. По нашим данным, признаки роевого состояния при отсутствии продуктивного медосбора наблюдались с 17 июня по 13 июля у 30% пчелиных семей полесской и уральской популяций и у 60% татарской. За этот период в пчелиных семьях было заложено в среднем по 10 маточников. С появлением медосбора с гречихи (2,0—2,5 кг в сутки) пчелиные семьи вышли из роевого состояния и переключились на сбор меда. При этом они уничтожили не только роевые мисочки, но и зрелые маточники.
Интересно отметить, что склонны к роению пчелиные семьи со старыми матками (3 года и старше). В год вывода матки и на следующий они практически не приходят в роевое состояние. Таким образом, использование молодых маток может быть хорошим приемом против роения у среднерусских пчел.
Матки среднерусских пчел отличаются хорошей плодовитостью, откладывая при благоприятных условиях до 2000 и более яиц в сутки. Среднесуточная плодовитость маток других пород не превышает у серой горной кавказской 1500 яиц, у карпатской—1800 яиц, у краинской—1400—2000 яиц. И только матки итальянской породы, способные откладывать более 2500 яиц в сутки, превосходят среднерусских по этому признаку.
Особенностью среднерусских пчел является усиленное выращивание расплода при слабом медосборе,когда пчелы южных пород, напротив, ограничивают яйцекладку маток.
Если есть возможность выбирать источник медосбора, среднерусские пчелы предпочитают такие медоносы, как гречиха, липа, и работают на них лучше пчел южных пород (В.П.Великанов, 1927; Н.В.Андреева, 1929).
Неоднократно отмечалось, что среднерусские пчелы хорошо собирают мед только при обильном медосборе и значительно отстают от серых горных кавказских при слабом полифлерном медосборе (например, клеверно-луговом) (Г.Д.Билаш, 1962; А.Я.Шекшуев, 1965).
Г.Д.Билаш (1966) считает, что у среднерусских пчел хорошо выражена флороспециализация к нектару гречихи, а также существует сильная привязанность к однажды выявленному источнику медосбора, что может обеспечивать при определенных условиях их преимущество над серыми горными кавказскими.
Некоторые исследователи объясняют различия в флоромиграции, пыльцесобирательной деятельности, использовании того или иного типа медосбора условиями, в которых шла эволюция пород пчел. Если южные породы формировались в условиях длительного, ровного и спокойного медосбора — с большим разнообразием растений с яркоокрашенными цветками, то становление среднерусской породы проходило в условиях лесного типа — с более узкой полифлерностью медосбора и менее яркой окраской цветков медоносов. В первом случае связи пчел с растениями были непрочны, старые рефлексы быстрее угасали, легко вырабатывались новые. В последнем случае пчелы (среднерусские) приспосабливались к короткому активному сезону с бурным медосбором, у них вырабатывалась консервативность в отношении флоромиграции (Н.А.Тушмалова, 1958).
Этим различиям могут быть и другие объяснения, но проведенные наблюдения показывают, что среднерусские пчелы редко мигрируют (Ш.О.Гасанов, 1970) и они лучше собирают пыльцу с бледноокрашенных цветков (Р.К.Миронова, 1974).
Среднерусские пчелы отличаются исключительно высокой зимостойкостью, не зная себе равных по этому признаку среди других пород. Кроме тoгo, они устойчивы к ряду заболеваний (нозематоз, падевый токсикоз).
Все это позволяет успешно разводить их в северных районах страны, где безоблетный период продолжается до 6 месяцев. В местах с сильным поздним медосбором среднерусские пчелы собирают значительные запасы меда.
Из других биологических особенностей среднерусской породы следует отметить следующие:
- чистопородные среднерусские пчелы — темноокрашенные, тергиты не имеют признаков желтизны, стерниты — от темно-серого до темно-коричневого цвета; конец брюшка у рабочих особей более тупой, чем у пчел других пород; посадка спокойно сидящей пчелы низкая, приземистая;
- при осмотре гнезд пчелы очень беспокойны и пугливы, сбегают с сотов вниз и свисают на нижней планке осматриваемой рамки рыхлыми, легко срывающимися гроздьями; злобливость отмечена во всех популяциях, особенно у вологодских и уральских пчел, проявление ее зависит от очень многих внешних факторов; окуривание дымом сильно снижает агрессивность пчел, однако злоупотребление дымом также может привести пчелиную семью в ярость (См.Особенности проявления агрессивности средне-русски...) ;
- при слабом поступлении нектара пчелы охотнее удлиняют медовые ячейки в верхних углах сота, чем отстраивают новые на противоположной стороне улочки; восковые перемычки в верхней части сотов и восковые мостики на печатке меда между двумя сотами отсутствуют либо они редкие, тонкие, лепестковой формы; глубина ячеек в расплодной части рамок всюду одинакова, печатка меда преимущественно белая, «сухая», а при очень слабом, неустойчивом медосборе иногда наблюдается переходный тип печатки;
- при слабом медосборе яйцекладка маток пчелами не ограничивается, выращивание расплода усиливается, а при сильном — яйцекладка резко сокращается и энергия пчел переключается на накопление запасов меда.
Еще Э.Бертран (по Л.Лангелю, 1903—1904) говорил: «Мы бы желали дать один хороший совет начинающим пчеловодам, а именно: не слишком быстро увлекаться иностранными породами пчел. Наша простая северная порода превосходна и во всех отношениях лучше всего пригодна для изучения на ней пчеловодства».
Популяции среднерусских пчел — исходный материал для селекции
Итак, пчелы среднерусской породы по целому комплексу признаков отличаются от пчел других пород, но и внутри породы они тоже неоднородны. Занимая огромный ареал, обитая зачастую в резко отличающихся друг от друга природно-климатических условиях, под влиянием естественного отбора и частично при воздействии человека среднерусская порода образовала обособившиеся группы, приспособленные к тем или иным конкретным условиям. Это характерно для всей породы, а не только для ее части, обитающей в пределах нашей страны.
Еще профессор Г.А.Кожевников (1929), ссылаясь на Е.Цандера, писал, что в Германии различали настоящую темную лесную пчелу, мало склонную к роению, и вересковую пчелу, обитающую на Северо-Западе страны и невероятно ройливую. Ф.Бельденспержер (1928) отмечал, что германские лесные пчелы во многом отличаются от французских. X.Н.Абрикосов (1946) сообщал о голландской вересковой темной пчеле, которая была завезена в Америку в 1638г. Она расселилась в дуплах и расщелинах скал. По наблюдениям В.В.Алпатова (1948), темная лесная пчела, завезенная в США из Голландии, отличается свирепым нравом, в то время как в Англии она сравнительно миролюбива.
Доминирующая на территории Франции темная лесная пчела представлена большим количеством экотипов, приспособленным к местным, узко локализованным, экологическим условиям. Но и в новой местности, в несвойственных им условиях, пчелы сохраняют прежний ритм развития. Таким образом, эта приспособленность наследственна (Ф.Руттнер, 1969; Ж.Луво, 1969).
В Польше издавна содержались такие популяции местных пчел, как среднеевропейская, вересковая и лесная (боровая) (/.Woznica, 1972).
М.Gromisz (1982) склонен выделять в пределах Apis mellifera mellifera такие популяции, как иберийская, среднеевропейская, вересковая, лесная, нигра.
Подобные группы пчел различными исследователями называются по-разному — экотипами, популяциями и даже породами. Именуются такие группы чаще всего по географическому принципу.
На территории бывшего СССР выделены некоторые своеобразные группы среднерусских пчел, которые, с известными допущениями и на уровне современных представлений, можно назвать сложившимися популяциями.
Башкирская (бурзянская) бортевая пчела. Обитает в лесах Бурзянского заповедника, сейчас широко известна. Один из корифеев пчеловодной науки профессор Г.А.Кожевников (1931) об этой пчеле писал так: «В бортях Башкирии и Уральской области мы имеем остаток этой первобытной европейской лесной темной пчелы, которая в настоящее время представляет с точки зрения генетики величайшую драгоценность. Ее надо всемерно беречь от метизации и на ней основать массовое разведение коренной среднеевропейской пчелы, которая тысячелетним отбором в жестокой борьбе за существование среди суровой горной природы выработала в себе выносливость и устойчивость».
Бурзянские пчелы отличаются рядом особенностей: средняя масса рабочей пчелы составляет 109 мг, нагрузка медового зобика —74—87 мг, длина хоботка — 6,10 мм. Пчелы выделяют много воска, дневной принос семьей пчел меда с липы может превышать 10 кг, а яйценоскость маток в июне иногда превышает 2500 яиц в сутки. Роение начинается с первой декады июня и обрывается только с наступлением главного медосбора, при этом закладывается 8—14 маточников. Нередко наблюдается «израивание» семей; такие семьи, как правило, не перезимовывают.
Бурзянские пчелы чутко реагируют на резкие изменения погодных условий и дружно возвращаются к жилью. Воровство наблюдается крайне редко. Одна из характерных особенностей этих пчел — сгрызание вощины в бортях.
В течение августа пчелы готовятся к суровой зиме: собирают пыльцу, мед, заклеивают все щели прополисом, уменьшают летки. Зимовка длится полгода, зимняя температура — минус 40—45°С. При первом весеннем облете бортевые пчелы уже несут пыльцу (В.Г.Генрих, В.А.Тюльпанова, 1958; A.Л.Дулькин, Г.Ф.Трескова, 1953; Е.М.Петров, 1983) (См. Башкирская пчела. , Бурзянские бортевые пчелы и варроатоз. , Заповедник Шульган - Таш и бурзянская бортевая пче...).
Уральская горно-таежная пчела. Исключительно хорошо приспособлена к условиям Северного Урала. Масса рабочей пчелы этой популяции составляет 88— 106 мг, восковая железа развита хорошо, нагрузка медового зобика — 51—72 мг, матка может откладывать до 1800 яиц в сутки. В роевом состоянии закладывается от двух до 28 маточников, чаще 5. Первые вылеты пчелы способны делать в конце февраля — начале марта, уже при температуре плюс 3—4°С. Пчелы могут летать в пасмурную, дождливую и ветреную погоду. Рабочий день на главном медосборе продолжается до 16 ч (В.Синцов, 1951; A.Л.Дулькин, 1958). (См. Средне-русская пчела на севере Урала.).
Алтайская пчела. С.Г.Миньков и другие (1969) приводят данные о появлении пчел в Сибири. Впервые на Алтай (в Усть-Каменогорскую крепость) 30 колод с пчелами были завезены в 1776 г. из Башкирии по ходатайству главного медика пограничных сибирских войск Беренса. Пчелиные семьи хорошо развились, дали по 2—3 роя и по 8 кг меда. Однако зимних кормов было оставлено в колодах недостаточно, и в период зимовки 1778 г. пчелы полностью отошли.
В 1786 г. по просьбе полковника Аршеневского пчелы были повторно завезены из Полесья Киевской губернии. Колоды доставлялись на подводах, и из 24 колод в 12 пчелы были живы. Условия оказались благоприятными для пчел: их стали разводить, а затем и продавать.
За пчелами приезжали отовсюду, и в скором времени вся Чулымская тайга, пространство староказацкой линии между Кузнецком и Бийском, стали базой развития пчеловодства в Сибири. Пчелы определенным образом приспособились к местным условиям, и Н.Ф.Крахотин (1960) отмечает у них ряд особенностей.
Алтайские пчелы не очень ройливы, но в отдельные годы в роевое состояние приходит до 90% семей, при этом закладывается от 5 до 16 маточников. Период роения длится 50—55 дней. Длина хоботка рабочих пчел составляет 6,2 мм, нагрузка медового зобика — 31,5 мг, максимум яйценоскости маток—1575 яиц в сутки. Одна из характерных особенностей этих пчел — встречающиеся при запечатывании меда на гранях крышечек выпуклости в виде жилок светло-желтого цвета, напоминающие по виду завитки шерсти.
Существуют и другие популяции, о которых упоминается в литературе,— алтайская горно-таежная (А.Л.Дулькин, 1960; Н.М.Ершов, 1970), березинская и полесская (В.М.Неручев, 1968; А.С.Гурченко, 1981) и др. Сведения о популяциях среднерусских пчел в Татарии сообщает Т.С.Жданова (1970), а в Мордовии - Т.А.Анциферова и Л.С.Швецова (1970).
С целью изучения генофонда среднерусской породы в 1970—1972 гг НИИ пчеловодства было проведено экспедиционное обследование наиболее типичных областей ее естественного ареала. Цель его — поиск и завоз сохранившихся в чистоте среднерусских пчел, а также выделение внутрипородных популяций и использование перспективного исходного материала для дальнейшей селекции и репродукции.
Располагая крайне скудной информацией по этому вопросу, мы сочли целесообразным подобрать и завезти пчел из мест с наибольшей географической удаленностью друг от друга. Обследовали пасеки, на которые в течение послевоенного периода не завозили пчелиных семей и маток других пород. Отбирая в полевых условиях пчелиные семьи, учитывали следующие признаки: окраску тела пчел и форму конца брюшка, позу пчел в состоянии покоя, активность защиты гнезда, реакцию на дым и поведение на вынутой из улья соторамке, характер отстройки сотов и форму их медовой и расплодной части, печатку меда, наличие и особенности восковых перемычек между сотами. Несмотря на условность каждого из перечисленных признаков, применение их в комплексе обеспечило надежный отбор чистопородного материала, что подтвердилось дальнейшим изучением экстерьера пчел. В результате этой работы были завезены 7 популяционных групп среднерусских пчел, 6 из которых (из Татарии, Новосибирской, Вологодской,Владимирской, Челябинской и Орловской областей) были поставлены на испытание по комплексу хозяйственно полезных признаков.
Экстерьерные признаки среднерусских пчел
Начало морфологическим исследованиям пчел было положено Г.А.Кожевниковым (1900). А.С.Михайлов (1924), В.В.Алпатов (1925) установили зависимость размеров хитиновых частей пчел, и прежде всего длины хоботка, от географической широты местности. В настоящее время экстерьерные характеристики наряду с окраской тела пчел широко применяют для идентификации отдельных географических пород внутри вида. Но выделение внутри вида только географических пород явно недостаточно.
В последние годы значительно возрос интерес к изучению более мелких таксономических единиц пчелы медоносной. Это связано с проблемой сохранения и необходимостью более интенсивного привлечения к использованию в селекционных целях локальных внутрипородных популяций пчел, обладающих ценным генофондом.
Однако выделение локальных популяций вызывает у зоологов определенные трудности. Э.Майр (1968, 1971) отмечает, что степень различий между популяциями бывает от почти полной идентичности до различий практически видового уровня. Границы изменчивости соседних популяций обычно перекрываются. Соседние популяции оказываются сходными в отношении одних признаков и различными в отношении других. Тем не менее локальные популяции слегка отличаются друг от друга генетически и фенотипически. Для установления этих различий обычно требуются очень точные измерения и статистические методы. Если локальные популяции пчелы медоносной, значительно удаленные друг от друга по широте, удается сравнительно легко различать по морфологическим признакам, то подобный анализ популяций из мест, мало различающихся по широте, затруднен. Поэтому перспективным методом, позволяющим анализировать как сходство, так и различие популяций, кроме морфологического анализа может быть сравнение изменчивости признаков. При этом сравнивают как абсолютные значения коэффициентов вариации (С0%) или среднеквадратичных отклонений (сг), так и построенные на их основе графики. Степень изменчивости признаков может, по современным представлениям, свидетельствовать о самых начальных сдвигах в изменении морфологии популяций. В некоторых работах на разных зоологических объектах показана возможность использования параметров изменчивости для изучения популяционной структуры вида (А.В.Яблоков, 1966; А.П.Разоренова, 1975; Ю.И.Новоженов, 1978).
Пчел, завезенных из Владимирской, Вологодской, Орловской, Новосибирской, Челябинской областей и Татарии, мы проанализировали по экстерьеру (табл. 1).
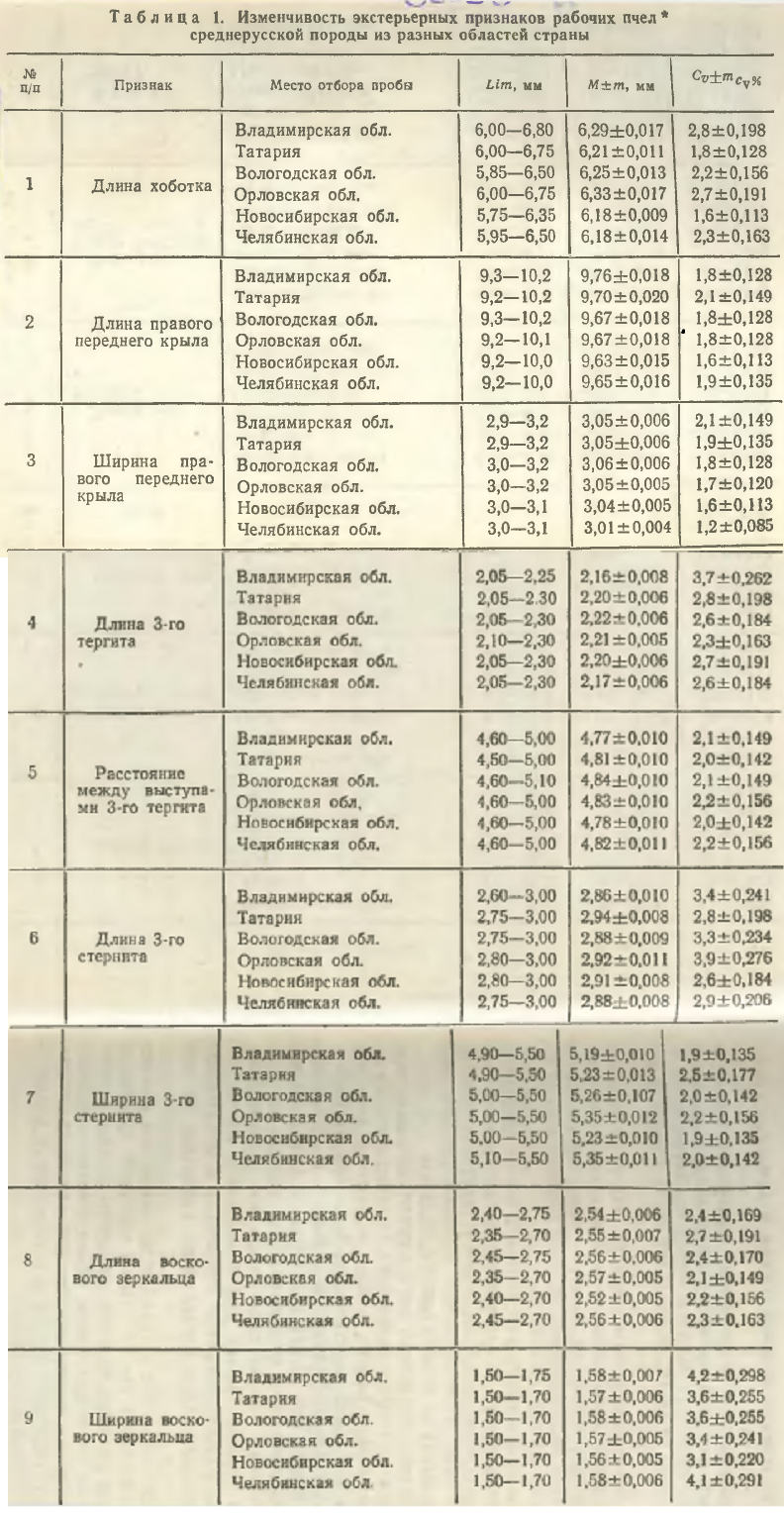 Между новосибирскими и челябинскими пчелами различий в длине хоботка не установлено. Наибольшие же расхождения по этому признаку (0,15 мм) обнаружены у пчел этих групп по сравнению с орловскими пчелами.
Между новосибирскими и челябинскими пчелами различий в длине хоботка не установлено. Наибольшие же расхождения по этому признаку (0,15 мм) обнаружены у пчел этих групп по сравнению с орловскими пчелами.Наибольшее значение средней длины крыла отмечено у владимирских пчел, а наименьшее — у новосибирских и челябинских (как и в предыдущем случае). Колебания размеров по этому признаку у всех сравниваемых групп пчел очень близки.
Наименьшая ширина крыла была у челябинских пчел, в остальных группах значения этого признака практически одинаковы и различия не превышают 0,02 мм. Колебания ширины крыла в пределах групп составляют 0,1—0,3 мм.
Наименьшее расстояние между выступами 3-го тергита (условная ширина) отмечено у владимирских и новосибирских пчел. Различия между крайними вариантами по этому признаку в пределах групп близки и составляют 0,4—0,5 мм.Наибольшая длина 3-го стернита зафиксирована у татарских пчел, наименьшая — у вологодских и челябинских, между которыми по этому признаку различия не оказалось. Максимальное значение длины 3-го стернита во всех группах одинаково.
Наибольшую ширину 3-го стернита имеют орловские пчелы, а наименьшую — владимирские. Одинаковые значения этого признака наблюдаются у татарских и новосибирских пчел. Максимальные значения ширины воскового зеркальца во всех группах одинаковы.
Хотя по всем признакам внутри групп отмечены «выбивающиеся» в сторону минимума и максимума значения, их, очевидно, можно отнести к разряду артефактов. Фенотипическая изменчивость этих признаков, судя по значениям коэффициента вариации (Сv%), сравнительно низка. Это может свидетельствовать о значительной однородности исследованного материала.
Проведенный анализ показывает, что внутри среднерусской породы (расы) можно выделить локальные популяции, обнаруживающие тенденцию к отклонению от средних значений в пределах ее стандарта по экстерьеру.
Так, новосибирские и челябинские пчелы по сравнению с остальными имеют самый короткий хоботок и наименьшие размеры правого переднего крыла, а между собой по этим признакам обнаруживают сходство.
Владимирские пчелы отличаются наибольшей длиной правого переднего крыла и наименьшими размерами 3-го тергита и 3-го стернита.
Татарские пчелы характеризуются наибольшей длиной 3-го стернита.
Орловские пчелы отличаются наибольшей длиной хоботка и наибольшими размерами 3-го стернита.
Отмеченные различия в размерах экстерьерных признаков изученных групп в большинстве случаев недостоверны и не могут в полной мере служить для объективного выделения локальных популяций.
Поэтому в дополнение к анализу абсолютных значений морфологических признаков мы проанализировали изменчивость экстерьерных признаков по значениям коэффициента вариации.
Проведенный анализ показал, что наибольшей изменчивостью характеризуются экстерьерные признаки владимирской популяционной группы, а наименьшей — новосибирской.
На рис. (с.22) показаны кривые величин коэффициента вариации экстерьерных признаков этих двух групп (прямая линия — владимирская, пунктир — новосибирская популяции), отражающие имеющиеся различия.
Наибольшую ширину 3-го стернита имеют орловские пчелы, а наименьшую — владимирские. Одинаковые значения этого признака наблюдаются у татарских и новосибирских пчел. Максимальные значения ширины воскового зеркальца во всех группах одинаковы.
Хотя по всем признакам внутри групп отмечены «выбивающиеся» в сторону минимума и максимума значения, их, очевидно, можно отнести к разряду артефактов. Фенотипическая изменчивость этих признаков, судя по значениям коэффициента вариации (Сv%), сравнительно низка. Это может свидетельствовать о значительной однородности исследованного материала.
Проведенный анализ показывает, что внутри среднерусской породы (расы) можно выделить локальные популяции, обнаруживающие тенденцию к отклонению от средних значений в пределах ее стандарта по экстерьеру.
Так, новосибирские и челябинские пчелы по сравнению с остальными имеют самый короткий хоботок и наименьшие размеры правого переднего крыла, а между собой по этим признакам обнаруживают сходство.
Владимирские пчелы отличаются наибольшей длиной правого переднего крыла и наименьшими размерами 3-го тергита и 3-го стернита.
Татарские пчелы характеризуются наибольшей длиной 3-го стернита.
Орловские пчелы отличаются наибольшей длиной хоботка и наибольшими размерами 3-го стернита.
Отмеченные различия в размерах экстерьерных признаков изученных групп в большинстве случаев недостоверны и не могут в полной мере служить для объективного выделения локальных популяций.
Поэтому в дополнение к анализу абсолютных значений морфологических признаков мы проанализировали изменчивость экстерьерных признаков по значениям коэффициента вариации.
Проведенный анализ показал, что наибольшей изменчивостью характеризуются экстерьерные признаки владимирской популяционной группы, а наименьшей — новосибирской.
На рис. (с.22) показаны кривые величин коэффициента вариации экстерьерных признаков этих двух групп (прямая линия — владимирская, пунктир — новосибирская популяции), отражающие имеющиеся различия.

Величины коэффициента вариации экстерьерных признаков татарской, вологодской и челябинской групп были близкими между собой и, как правило, занимали среднее положение по отношению к таковым владимирской и новосибирской групп пчел.
Анализируя полученные данные, мы видим, что Cv% владимирской популяции по всем признакам (кроме 7-го, по которому отмечены равные значения) больше новосибирской. Для 3, 4, 6, 9-го признаков Р>>0,99, а для первого признака Р>-0,90.
------------------------
Кривые величии коэффициента вариации экстерьерных признаков пчел этих групп на рис. не приводятся, так как они в значительной мере совпадают и занимают промежуточное положение между двумя кривыми, в некоторых случаях пересекая их. Характер изменчивости всех признаков сходен во всех выборках. Название признаков см. в табл. 1.— Примеч. авт.
Анализируя полученные данные, мы видим, что Cv% владимирской популяции по всем признакам (кроме 7-го, по которому отмечены равные значения) больше новосибирской. Для 3, 4, 6, 9-го признаков Р>>0,99, а для первого признака Р>-0,90.
------------------------
Кривые величии коэффициента вариации экстерьерных признаков пчел этих групп на рис. не приводятся, так как они в значительной мере совпадают и занимают промежуточное положение между двумя кривыми, в некоторых случаях пересекая их. Характер изменчивости всех признаков сходен во всех выборках. Название признаков см. в табл. 1.— Примеч. авт.
Таким образом, анализ изменчивости экстерьерных признаков шести групп пчел среднерусской породы показывает определенную специфичность новосибирской и владимирской популяционных групп. Татарские, орловские и вологодские пчелы, близкие по величине изменчивости, относятся, очевидно, к одной популяции, приближаясь по некоторым признакам или к владимирской, или к новосибирской популяции. Челябинские пчелы, отличаясь некоторой морфологической специфичностью, имеют в то же время признаки переходного характера, приближающие их, с одной стороны, к татарско-орловско-вологодской популяции, с другой— к новосибирской.
Коэффициент вариации как мерило изменчивости может быть одним из важных популяционных параметров, который целесообразно использовать при изучении популяционной структуры пчелы медоносной, особенно в тех случаях, когда не имеется четких морфологических различий.
С учетом очень высоких требований к точности и достоверности возникает вопрос, сколько пчел необходимо для характеристики экстерьера всей пчелиной семьи?
По Е.К.Меркурьевой (1970) определим количество пчел в пробе для получения достоверных данных при f0 999=з 291 и ошибке Е=2%. По предварительным расчетам установим, что мерные экстерьерные признаки имеют Cv на уровне 3%. Расчет по формуле показывает:

Таким образом, при очень высоких требованиях точности и достоверности проба из 24 рабочих пчел вполне объективно и достоверно характеризует экстерьер всей пчелиной семьи, и препарировать 50 пчел, как это часто рекомендуют, нет необходимости.
Хозяйственно полезные признаки пчелиных семей различных популяций
В 1972—1973 гг. каждую из завезенных популяционных групп размножали «в себе» в условиях пространственной изоляции и при наличии собственного мощного трутневого фона.
Для контроля мы взяли популяционную группу местных пчел Орловской области.
Зимы в центральных областях России нередко выдаются суровыми и продолжительными. Это показали и результаты зимовки популяционных групп среднерусских пчел в 1974—1976 гг. Оба года пчелы зимовали в полуподземном зимовнике. Крайне неблагоприятной для пчел была зима 1975/76 г., когда температура с 5 по 12 февраля опускалась до минус 35—38 °С. Пчел из зимовника выставили только 5—10 апреля. К этому следует добавить, что после зимовки 1974/75 г. во второй половине июля, а также в августе на Орловщине, как и в других смежных областях, медосбора полностью не было. Это в значительной степени затрудняло осеннее наращивание молодых пчел. И все же отхода пчелиных семей в период зимовки на пасеке-испытательнице практически не было, а зимний отход маток за две зимы в семьях местных орловских пчел составил всего лишь 2,5%, у вологодских пчел — 6,0, у уральских — 5,9, у новосибирских — 3,1, у владимирских — 4,4%. В пчелиных семьях татарской популяционной группы зимнего отхода маток вообще не отмечалось. Опоношенность в гнездах наблюдалась в основном после зимовки 1975/76 г., а в среднем за две зимовки она составила в татарской популяционной группе 0,88 балла, во владимирской—1,98, в новосибирской — 2,15, в уральской— 2,03, в вологодской— 1,29, в местной— 1,76 балла (опоношенность оценивали по пятибалльной шкале).
Интересно отметить, что у 62,5% пчелиных семей татарской популяции поноса не отмечалось совсем; без признаков опоношенности во владимирской популяционной группе зимовало 32% семей, в новосибирской — 18,4, в уральской — 33,0, в вологодской — 35,9, в местной— 32,5%. Основные результаты зимовки опытных пчелиных семей приведены в табл. 2.
Коэффициент вариации как мерило изменчивости может быть одним из важных популяционных параметров, который целесообразно использовать при изучении популяционной структуры пчелы медоносной, особенно в тех случаях, когда не имеется четких морфологических различий.
С учетом очень высоких требований к точности и достоверности возникает вопрос, сколько пчел необходимо для характеристики экстерьера всей пчелиной семьи?
По Е.К.Меркурьевой (1970) определим количество пчел в пробе для получения достоверных данных при f0 999=з 291 и ошибке Е=2%. По предварительным расчетам установим, что мерные экстерьерные признаки имеют Cv на уровне 3%. Расчет по формуле показывает:

Таким образом, при очень высоких требованиях точности и достоверности проба из 24 рабочих пчел вполне объективно и достоверно характеризует экстерьер всей пчелиной семьи, и препарировать 50 пчел, как это часто рекомендуют, нет необходимости.
Хозяйственно полезные признаки пчелиных семей различных популяций
В 1972—1973 гг. каждую из завезенных популяционных групп размножали «в себе» в условиях пространственной изоляции и при наличии собственного мощного трутневого фона.
Для контроля мы взяли популяционную группу местных пчел Орловской области.
Зимы в центральных областях России нередко выдаются суровыми и продолжительными. Это показали и результаты зимовки популяционных групп среднерусских пчел в 1974—1976 гг. Оба года пчелы зимовали в полуподземном зимовнике. Крайне неблагоприятной для пчел была зима 1975/76 г., когда температура с 5 по 12 февраля опускалась до минус 35—38 °С. Пчел из зимовника выставили только 5—10 апреля. К этому следует добавить, что после зимовки 1974/75 г. во второй половине июля, а также в августе на Орловщине, как и в других смежных областях, медосбора полностью не было. Это в значительной степени затрудняло осеннее наращивание молодых пчел. И все же отхода пчелиных семей в период зимовки на пасеке-испытательнице практически не было, а зимний отход маток за две зимы в семьях местных орловских пчел составил всего лишь 2,5%, у вологодских пчел — 6,0, у уральских — 5,9, у новосибирских — 3,1, у владимирских — 4,4%. В пчелиных семьях татарской популяционной группы зимнего отхода маток вообще не отмечалось. Опоношенность в гнездах наблюдалась в основном после зимовки 1975/76 г., а в среднем за две зимовки она составила в татарской популяционной группе 0,88 балла, во владимирской—1,98, в новосибирской — 2,15, в уральской— 2,03, в вологодской— 1,29, в местной— 1,76 балла (опоношенность оценивали по пятибалльной шкале).
Интересно отметить, что у 62,5% пчелиных семей татарской популяции поноса не отмечалось совсем; без признаков опоношенности во владимирской популяционной группе зимовало 32% семей, в новосибирской — 18,4, в уральской — 33,0, в вологодской — 35,9, в местной— 32,5%. Основные результаты зимовки опытных пчелиных семей приведены в табл. 2.

Анализ приведенных данных показывает, что наиболее сильными (7 улочек) из зимовки выходили семьи татарской популяции, превосходившие контроль по силе на 18,6%; на втором месте оказались семьи владимирской и новосибирской популяционных групп, на 8,5% уступающие по силе контролю. На первом месте по количеству печатного расплода во время 1-го весеннего осмотра оказались также пчелиные семьи татарской популяционной группы, на втором — семьи местных (орловских) пчел, на третьем — семьи вологодской и новосибирской популяционных групп. Тяжелая зимовка 1975/76 г. сильнее всего отразилась на зимнем отходе (осыпи) пчел, но ее влияние сказалось по-разному на пчелах опытных групп.Усредненные (за две зимовки) данные показывают, что менее всего пчел зимой осыпалось в семьях татарской популяционной группы (21,2%), на втором месте по-прежнему были пчелиные семьи местной популяционной группы, на третьем — вологодской.
На пасеке встречались также пчелиные семьи, в которых зимней осыпи практически не наблюдалось. Процент таких семей в татарской популяционной группе составил 23,5, во владимирской—17,0, в новосибирской— 15,5, в уральской — 6,5, в вологодской—11,0, в местной—11,7. Такие семьи представляют большой интерес для селекции по этому признаку.
Расход корма в пересчете на улочку зимовавших пчел был сравнительно небольшим (1,56— 1,65 кг), и разница между группами по этому показателю не превышала 0 — 7%.
В 1974—1975гг. испытывались также опытные группы пчелиных семей по яйценоскости маток, силе, медопродуктивности и воскопродуктивности в условиях лесостепи Орловской области. Ранневесенний медосбор на Орловщине обеспечивается обычно ивовыми и кленами. Весенний медосбор с косточковых садовых культур используется не всегда из-за обычных в это время похолоданий, а цветение семечковых (яблони, груши), как правило, совпадает с устойчивой теплой погодой. Из других весенних и раннелетних медоносов следует отметить желтую акацию, одуванчик лекарственный, а также клевера и сурепку. Из-за небольших площадей лугов в июне, и особенно во второй его половине, практически полностью отсутствует медосбор, и пчелиные семьи входят в роевое состояние. В среднем за два сезона по всей пасеке-испытательнице приходило в роевое состояние 24,8% семей, а роилось всего лишь 4,5%. Наибольший процент отроившихся семей (8,7) отмечен в татарской популяционной группе, в вологодской группе роившихся семей не было. Продуктивный медосбор начинается с конца июня с семенников гибридного клевера и различных сорняков.
При суточных привесах контрольного улья 2—3 кг семьи среднерусских пчел выходят из роевого состояния. При этом пчелы уничтожают даже запечатанные роевые маточники. В начале июля зацветает гречиха, обеспечивающая при благоприятных условиях главный медосбор.
Однако не только зимы, но и целиком сезоны 1974—1975 гг. оказались крайне неблагоприятными для жизнедеятельности пчелиных семей и их продуктивности. Так, в мае — июле 1974г. осадков выпало в 3—5 раз больше среднего многолетнего уровня, а лето было прохладным. Сезон 1975г. характеризовался, наоборот, сильнейшей засухой, которая резко снизила нектаропродуктивность лугового и полевого разнотравья и привела к полному отсутствию медосбора с гречихи.
Сложившиеся условия отрицательно влияли на яйценоскость маток, однако между популяционными группами наблюдались различия по этому признаку (табл. 3).
На пасеке встречались также пчелиные семьи, в которых зимней осыпи практически не наблюдалось. Процент таких семей в татарской популяционной группе составил 23,5, во владимирской—17,0, в новосибирской— 15,5, в уральской — 6,5, в вологодской—11,0, в местной—11,7. Такие семьи представляют большой интерес для селекции по этому признаку.
Расход корма в пересчете на улочку зимовавших пчел был сравнительно небольшим (1,56— 1,65 кг), и разница между группами по этому показателю не превышала 0 — 7%.
В 1974—1975гг. испытывались также опытные группы пчелиных семей по яйценоскости маток, силе, медопродуктивности и воскопродуктивности в условиях лесостепи Орловской области. Ранневесенний медосбор на Орловщине обеспечивается обычно ивовыми и кленами. Весенний медосбор с косточковых садовых культур используется не всегда из-за обычных в это время похолоданий, а цветение семечковых (яблони, груши), как правило, совпадает с устойчивой теплой погодой. Из других весенних и раннелетних медоносов следует отметить желтую акацию, одуванчик лекарственный, а также клевера и сурепку. Из-за небольших площадей лугов в июне, и особенно во второй его половине, практически полностью отсутствует медосбор, и пчелиные семьи входят в роевое состояние. В среднем за два сезона по всей пасеке-испытательнице приходило в роевое состояние 24,8% семей, а роилось всего лишь 4,5%. Наибольший процент отроившихся семей (8,7) отмечен в татарской популяционной группе, в вологодской группе роившихся семей не было. Продуктивный медосбор начинается с конца июня с семенников гибридного клевера и различных сорняков.
При суточных привесах контрольного улья 2—3 кг семьи среднерусских пчел выходят из роевого состояния. При этом пчелы уничтожают даже запечатанные роевые маточники. В начале июля зацветает гречиха, обеспечивающая при благоприятных условиях главный медосбор.
Однако не только зимы, но и целиком сезоны 1974—1975 гг. оказались крайне неблагоприятными для жизнедеятельности пчелиных семей и их продуктивности. Так, в мае — июле 1974г. осадков выпало в 3—5 раз больше среднего многолетнего уровня, а лето было прохладным. Сезон 1975г. характеризовался, наоборот, сильнейшей засухой, которая резко снизила нектаропродуктивность лугового и полевого разнотравья и привела к полному отсутствию медосбора с гречихи.
Сложившиеся условия отрицательно влияли на яйценоскость маток, однако между популяционными группами наблюдались различия по этому признаку (табл. 3).

Яйценоскость маток определяет такой важнейший хозяйственно полезный признак пчелиных семей, как их сила.
Пчелиные семьи татарской популяции по силе на медосборе превышали контроль на 17,5%, на втором месте по этому признаку стояли пчелиные семьи местной и вологодской популяционных групп. Владимирская, новосибирская и уральская популяционные группы были равными по силе и на 0,2 кг уступали контролю.
Различные темпы развития пчелиных семей опытных групп, разная сила на медосборе в значительной мере способствовали их различной продуктивности (табл. 4).

Крайне неблагоприятные погодные условия сезонов 1974—1975 гг. привели к низким медосборам в целом по Орловской области. Так, в Орловском и Урицком районах (в местах расположения кочевых точков пасеки-испытательницы) средний медосбор общественных пасек в 1975 г. составил, соответственно, 14,4 и 13,4 кг
Медопродуктивность пасеки-испытательницы в целом оказалась выше. Наилучшими показателями отличались семьи татарской популяционной группы, на 25,7% по меду и на 13,6% по воску превзошедшие контроль. Превосходство продуктивности татарских пчел над остальными опытными группами было еще большим. Второе и третье места разделили по этому признаку пчелиные семьи вологодской и местной популяционных групп.
Сравнительное испытание шести популяционных групп среднерусских пчел в условиях Орловской области позволило установить некоторые различия между ними. Наилучшей зимостойкостью и наиболее высокой яйценоскостью маток, силой и продуктивностью пчелиных семей отличается татарская популяционная группа, на втором месте оказалась местная (орловская) популяционная группа пчел и на третьем — вологодская.
Во всех изученных популяционных группах среднерусских пчел наблюдается значительная фенотипическая изменчивость хозяйственно полезных признаков,которая может быть предпосылкой эффективной селекции по ним.
Приведенные данные конечно же не отражают все популяции и специфические группировки среднерусских пчел, но они иллюстрируют определенные особенности этих групп и свидетельствуют о значительных генетических резервах среднерусской породы. Учитывая особенности среднерусских пчел, селекционная программа по улучшению породы должна поддерживать имеющийся высокий уровень зимостойкости и развивать потенциальные возможности эффективного использования бурного, но короткого медосбора средней полосы.
ТЕОРЕТИЧЕСКИЕ ОСНОВЫ СЕЛЕКЦИИ СРЕДНЕРУССКИХ ПЧЕЛ
Селектируемые признаки: наследуемость, изменчивость, повторяемость
Подавляющее большинство хозяйственно полезных признаков сельскохозяйственных животных (в том числе и пчел) относится к категории количественных, они контролируются многими генами и детерминируются факторами внешней среды. Наследование полигенных признаков и другие генетико-популяционные параметры современная генетика изучает не на отдельных особях, а в популяциях, с помощью различных математико-статистических методов.
Для генетической характеристики популяций используют такие параметры популяционной генетики, как их фенотипическая и генотипическая изменчивость, наследуемость, повторяемость, фенотипическая и генотипическая корреляция.
По мнению Ф.Ф.Эйснера (1981), эффективность селекции обусловлена величиной селекционного дифференциала, степенью наследуемости селектируемых признаков, быстротой смены поколений, числом признаков, по которым ведут селекцию, характером коррелятивных связей между ними.
Наследуемость позволяет прежде всего судить о генотипическом разнообразии признака, а также дает возможность прогноза селекционной ценности особи по ее фенотипическим проявлениям уже на первых этапах селекционного процесса.
Показатели наследуемости (h2-H2) служат одним из параметров при перспективном планировании селекции. В то же время в каждом массиве животных необходим самостоятельный анализ коэффициентов наследуемости (В.Н.Новоставский, 1969).
Показатель наследуемости (Л2*//2) позволяет точнее установить степень сходства между племенными животными и их потомством, что и определяет его практическое значение в селекции. При более высоком значении h2 отбор по фенотипу лучших особей создает более широкие возможности получения лучшего потомства, а при низких значениях № отбор малоэффективен.
Первые расчеты показателя наследуемости в пчеловодстве касались медопродуктивности (F.Pirchner, F.Ruttner, Н.Ruttner, 1960). Было показано, что № медопродуктивности составляет в среднем 0,2—0,3, что соответствует значениям такового для продуктивности других сельскохозяйственных животных.
Затем исследователи стали принимать попытки использования показателя наследуемости для прогноза селекции на одно поколение (В.Веселый, Р.Шиллер, 1963; М.А.Ел-Бамби, 1965; М.Gromisz, 1972; 1978).
Zawilski (1972) определил коэффициенты наследуемости некоторых экстерьерных признаков пчел. Они оказались выше, чем для показателей продуктивности (h2 = 0,40 — 0,75).
Т.Е.Rinderer, A.М.Collins, М.A.Brown (1983) в Батон-Руже (США) определили h2 устойчивости к нозематозу, продолжительности жизни, реакции на феромон тревоги изопентилацетат. Показатели наследуемости продолжительности жизни пчел и реакции на изопентилацетат составили 0,30—0,32, остальные коэффициенты были недостоверны.
R.F.Moritz (1985) в ФРГ определил коэффициенты наследуемости стадии печатного расплода (h2 = 0,8) и попытался связать продолжительность стадии печатного расплода с устойчивостью к варроатозу. Показатель наследуемости продолжительности жизни пчел у Е.P.Milne (1985) оказался значительно ниже (h2 = 0,196).
Этот же автор изучил наследуемость и целого ряда других признаков; кормодобывающего поведения (на основе лабораторных тестов), массы куколки и т. д.
Приведенные выше сведения показывают, что проблеме наследуемости признаков пчел во всем мире уделяется серьезное внимание. Однако величины показателей наследуемости одних и тех же признаков у разных авторов не совпадают. Это объясняется различными способами расчета показателя h2, а также влиянием конкретных условий. Одна из особенностей h2 заключается в том, что его значение изменяется с изменением условий, зависит от уровня и продолжительности селекционной работы и т. д.
Таким образом, анализ наследуемости признаков необходимо проводить в каждом конкретном случае самостоятельно одним из апробированных методов.
Если наследуемость имеет место при передаче генетической информации от поколения к поколению, то повторяемость (г, г2), по определению Н.А.Плохинского (1969), возникает при реализации одной и той же групповой генетической информации в разных возрастах и в разных условиях: лучшие в раннем возрасте не всегда остаются лучшими в последующих возрастах, лучшие в одних условиях жизни могут стать средними и даже худшими в той же группе, перешедшей в другие условия. Таким образом, повторяемость — это большее или меньшее постоянство рангов особей по их фенотипу в одной и той же группе, но в разных возрастах и в разных условиях жизни.
При высокой повторяемости признака становится эффективным и отбор по нему в раннем возрасте или в любых условиях. В определенном смысле показатель повторяемости служит показателем эффективности раннего отбора по фенотипу или отбора в любых условиях.
Как в начале селекционного процесса, так и в различных его стадиях необходимо иметь данные об изменчивости селективного признака. Чаще всего используется фенотипический коэффициент вариации (Vp)t однако более информативен генотипический коэффициент вариации (Vg), вычисляемый в результате дисперсионного анализа. К сожалению, пока что этот коэффициент селекционерами используется крайне редко (В.А.Мошкин, И.К.Сачли, 1977).
В.П.Коваленко (1976) сделал попытку использовать для выбора методов селекции и оценки ее эффективности на примере яйценоскости кур соотношение показателей изменчивости, наследуемости и продуктивности. Н.А.Соболев (1972) предложил применять в селекционной практике такие параметры, как относительная наследственная стабильность (g2) и вероятность отбора генотипов по фенотипу (р).
В пчеловодстве в той или иной мере используются лишь данные о степени наследуемости признаков и их фенотипической изменчивости. К тому же эти параметры вычисляются различными способами, что приводит к разногласиям между отдельными авторами. Поэтому одной из задач, стоявших перед нами, было подобрать подходящие методы для расчета этих параметров с целью их возможного использования в селекции.
Данные по некоторым генетико-популяционным параметрам среднерусских пчел приведены в табл. 5.

Коэффициенты фенотипической вариации за разные годы по одним и тем же признакам отличаются незначительно.
Коэффициент генотипической вариации (Vg) используется селекционерами редко, однако он более информативен, так как позволяет судить о гетерогенности по данному признаку и возможностях отбора в популяции. По всем изученным признакам в среднем Vg меньше Vp примерно в 3,4 раза. Нулевых значений Vg, свидетельствующих об отсутствии возможностей отбора, не зафиксировано ни по одному признаку. Колебания Vg по годам объясняются, очевидно, усилением взаимодействия генотипа и среды в неблагоприятные по погодным и медосборным условиям сезоны.
Значения коэффициентов наследуемости, рассчитанные двумя способами, практически не различались. Разница в сторону снижения коэффициентов наследуемости, рассчитанных с разложением факториального среднего квадрата, наблюдалась лишь при ничтожных «0,1 значениях Н2.
Сравнительно более высокие коэффициенты наследуемости отмечены для максимальной яйценоскости маток и для зимостойкости, выраженной суммой баллов. В первом случае превышение Н2 максимальной яйценоскости над среднесуточной можно объяснить тем, что в период проявления максимальной яйценоскости (конец июня) создаются наиболее благоприятные условия для жизнедеятельности пчелиных семей и работы маток. Во втором случае зимостойкость, выраженная суммой баллов, содержит гораздо более полную информацию о способности пчелиных семей переносить неблагоприятные зимние условия, чем отдельные показатели (отход пчел за зиму, количество печатного расплода весной, активность каталазы ректальных желез), и этот коэффициент наследуемости (Я2) тоже выше.
Значения коэффициентов наследуемости тем точнее чем больше имеется информации об изучаемом признаке. Коэффициенты наследуемости признаков, рассчитанные по усредненным за два года данным, выше рассчитанных по однолетним данным. Так, Н2 медопродуктивности при таком расчете составляет 0,27, среднесуточной яйценоскости маток— 0,24, максимальной яйценоскости 0,31.
Нестабильность значений Н2 может зависеть от многих причин, и в том числе от факторов среды, уровня продуктивности и возраста пчелиных маток (например, в 1972 г. матки были однолетними, а в 1973 г. — двухлетними) .
Высокие значения коэффициентов наследуемости указывают на эффективность отбора по селективным признакам. Однако Н.А.Соболев считает, что, получив низкие значения показателей наследуемости, нельзя считать бесполезным отбор по данному признаку, так как возможна высокая относительная наследственная стабильность признака (g2), если уровень общей вариации невысок. За исключением отдельных показателей зимостойкости (отход пчел за зиму, количество печатного расплода во время первого весеннего осмотра), в наших опытах наблюдалась довольно высокая относительная наследственная стабильность хозяйственно полезных признаков пчел (0,898—0,982).
Высокой относительной наследственной стабильности соответствует и обнадеживающая величина вероятности отбора генотипов по фенотипу (Р), позволяющая предполагать, что для большинства хозяйственно полезных признаков на правильный отбор хороших генотипов можно рассчитывать примерно в восьми случаях из десяти отобранных по фенотипу семей. Для количества печатного расплода во время 1-го весеннего осмотра такой отбор малоэффективен (Р = 0,6340—0,6558), а для отхода пчел за зиму нецелесообразен (Р<0,5000).
Результаты этих исследований позволяют считать, что изученные признаки отличаются большой фенотипической изменчивостью. Коэффициенты генотипической изменчивости ниже фенотипических, однако уровень генотипической гетерогенности достаточен для проведения отбора. Наиболее низкие значения коэффициентов наследуемости, а также относительной наследственной стабильности и вероятности отбора генотипов по фенотипу наблюдаются у признаков, характеризующих зимостойкость. При селекции по этим признакам массовый отбор будет, очевидно, неэффективен, и для их улучшения необходим более высокий уровень селекционной работы.
Коэффициенты наследуемости признаков, характеризующих продуктивность и плодовитость пчел, имеют значения на уровне таковых и для других сельскохозяйственных животных. Расчеты показывают, что при селекции по этим признакам из десяти отобранных по фенотипу пчелиных семей мы можем надеяться на правильный отбор восьми хороших генотипов.
Коэффициенты наследуемости таких признаков, как масса маток, длина хоботка, приближаются по своим значениям к единице. Они лишь в ничтожной степени зависят от паратипических факторов, а коэффициент наследуемости такого признака, как склонность к сбору гречишной пыльцы, составляет всего лишь 0,02.
Один из важнейших хозяйственно полезных признаков — среднесуточная яйценоскость маток. За три года повторяемость среднесуточной яйценоскости среднерусских маток составила:

факторов в разные периоды сезона составила в первые два года 26 - 29%, а в 1973 г.— 57% за счет выравненных условий последнего сезона. При этом возникает возможность объективной оценки признака по первым единичным учетам.
Как уже было отмечено, генетико-популяционные параметры селективных признаков динамичны. Мы проследили их изменение на протяжении восьми поколений отбора. Повторяемость определяли как коэффициент корреляции по одному признаку за два смежных года:

Таким образом, генотипическая изменчивость основных селективных признаков пчел имеет низкие или средние значения (6—34%). То же относится и к степени наследуемости этих признаков. Отсюда можно сделать предварительные выводы: на пасеках с низкой медопродуктивностью массовый отбор вряд ли будет достаточно эффективен. Очевидно, более перспективным окажется индивидуальный отбор с проверкой маток по качеству потомства. Если же уровень селективного признака достаточно высок (например, яйценоскость маток), может быть перспективна дальнейшая селекция по линиям.
Поскольку уровень наследуемости, изменчивости, повторяемости изменяется, необходим постоянный контроль этих генетико-популяционных параметров с целью корректировки селекционного процесса.
Фенотипическая корреляция
Основы учения о корреляции заложены еще Ч.Дарвином и Ж.Кювье. Ч.Дарвин (1859) утверждал, что если человек упорно ведет отбор по какому-либо определенному признаку и, следовательно, все более изменяет его, то в силу таинственных законов корреляции ом почти наверняка вызовет изменения и других частей организма.
Ж.Кювье (по И.И.Шмальгаузену, 1969) писал: «Так как все органы животного образуют единую систему, части которой зависят друг от друга и действуют и противодействуют одна по отношению к другой, никакое изменение не может обнаружиться в одной части без того, чтобы не вызвать соответствующего изменения во всех остальных частях».
И.И.Шмальгаузен (1969), возвращаясь к этой проблеме, отмечает, что различные формы корреляции являются выражением целостности организма в его индивидуальном развитии, которая проявляется в регулярных его реакциях на изменения внешней среды, и даже в реакциях на некоторые нарушения во внутреннем его строении. В процессе эволюции происходит очень медленное, но постоянное накопление корреляционных механизмов общего значения.
В настоящее время установлены корреляции различных признаков. Прежде всего, это корреляция между силой пчелиной семьи и ее продуктивностью. Еще А.С.Михайлов (1927), подтверждая старую истину: «В сильных во время взятка семьях — все спасение», установил достоверную и высокую корреляцию между этими признаками (г = 0,74). Достоверной связи продуктивности с весом пчел и некоторыми экстерьерными признаками он не обнаружил.
В. С. Коптев (1965) показал взаимосвязь между силой семей и медосбором (г = 0,55 — 0,84); G.Н.Cale (1968) установил связь между яйцекладкой матки, сбором пыльцы и медосбором (г = 0,50 — 0,81). Подобную зависимость подтверждают К.Давыденко (1970), Ю.А.Субботин (1971), V.Cavoysky (1977), D.Z.Nelsen, N.Е.Gary (1983). U.Smely (1984), основываясь на высокой корреляции медопродуктивности и площади расплода в начале цветения яблони, предложил использовать последний показатель для прогноза медосбора. Высокую корреляцию между медопродуктивностью и количеством расплода подтверждает I.Woyke (1984).
Предпринимаются попытки выявления корреляций между экстерьерными и интерьерными признаками пчел и маток и медопродуктивностью. Еще В.О.Пикель (1926) писал о зависимости медопродуктивности от длины хоботка и размеров тела пчел, а С.Г.Петров (1927) пытался прогнозировать медопродуктивность по развитию большой ядовитой железы. Однако А.Велих (1929) отрицал связь между длиной большой ядовитой железы, злобливостью и медопродуктивностью.
А.Н.Колесников (1959) установил прямую корреляцию между продуктивностью и такими признаками, как длина крыла, количество зацепок, длина и ширина воснового зеркальца и особенно ширина первого членика задней лапки, размеры крыла и тергитов.
I.Heitmanek (1960) отрицал влияние длины хоботка рабочих пчел на медосбор. Вместе с тем М.Н.Виноградов, В.М.Фролова (1969) убедительно показали на примере серых горных кавказских пчел влияние целого ряда экстерьерных признаков, и прежде всего длины хоботка, на медопродуктивность. В.Г.Кашковский (1965) считал, что для Кемеровской области более подходят пчелы с коротким хоботком.
С.P.Milne (1980) выявил связь веса куколок на стадии розовых глаз с медопродуктивностью. Г.А.Аветисян (1961) установил значительную корреляцию между количеством яйцевых трубочек, массой матки и ее плодовитостью, а Тхай-Бин-Ван (1975) — массой маток, количеством яйцевых трубочек и суммой длин 2—5-го тергитов. А.Н.Мельниченко и др. (1968),А.С.Тришина, Н.Д.Шмелева (1974) отрицали наличие какой-либо связи между яйценоскостью маток, количеством и длиной яйцевых трубочек, размерами маточников и массой маток.
Однако многие авторы даже рассчитывали коэффициенты корреляции между интерьерными и экстерьерными признаками. А.С.Михайлов (1927) установил связь между весом рождающейся пчелы и ее размерами, а также между длиной переднего крыла и числом зацепок на заднем крыле у тульских пчел. Т.Szabo (1973) нашел прямую зависимость между весом народившихся маток и их весом после прекращения яйцекладки.
Обнаружена корреляционная зависимость таких признаков, как площадь воскового зеркальца и сумма ширин 3-го и 4-го тергитов (L.Bornus, 1960), длина 3-го тергита и общая длина тела (В.Губин, 1969).
В. В. Тряско (1961), Г.Д.Билаш, В.Т.Желтякова (1965) рассчитали целый комплекс корреляционных связей между экстерьерными признаками.
В.В.Алпатов (1927) установил связь между силой пчелиной семьи и весом выводящихся в ней пчел (г= = 0,489), но А.С.Михайлов (1927) эту зависимость не
подтвердил.
М.В.Жеребкин (1969) выявил довольно высокую корреляцию между активностью инвертазы глоточных желез и медопродуктивностью (г=0,51— 0,87) и предложил использовать этот признак для прогнозирования медопродуктивности.
Т.Szabo (1974) предположил прямую взаимосвязь между массой матки на выходе и в период яйцекладки.
С.Mraz (1981) считает, что агрессивность пчел часто связана с их высокой жизнестойкостью, резистентностью к заболеваниям и высокой медопродуктивностью.
Таким образом, селекционеры предпринимают попытки расчета корреляций между самыми различными признаками пчел. Одна из целей этой работы — облегчить отбор признаков, так как селекция по целому комплексу параметров менее эффективна, но более трудоемка. При установлении же положительной корреляции возможен косвенный отбор одних признаков по другим, более доступным для учета. Изучение корреляций позволяет проводить раннюю оценку, прогнозировать развитие селективных признаков. Правда, мнения специалистов о существовании корреляции между одними и теми же признаками противоречивы, а то и диаметрально противоположны. Но они лишь подтверждают положение о том, что коэффициенты корреляции имеют смысл лишь в определенных условиях и на конкретных объектах, и в каждом отдельном случае их следует рассчитывать самостоятельно.
Селекция пчел по таким признакам, как медопродуктивность и зимостойкость пчелиных семей, яйценоскость маток, как, впрочем, и по большинству хозяйственно полезных признаков других сельскохозяйственных животных, затруднена вследствие того, что эти признаки контролируются многими генами и в значительной степени детерминируются факторами внешней среды.
Относительно высокое влияние этих факторов на коэффициенты наследуемости хозяйственно полезных признаков отмечают многие исследователи независимо от различий в используемых методиках расчетов (Н.А.Плохинский, 1964, П.Рокицкий, 1978).
В данной ситуации нельзя пренебрегать возможностями косвенного отбора пчел на медопродуктивность, зимостойкость, яйценоскость маток, используя коррелятивные связи.
Эффективность селекции, как уже было сказано, зависит как от степени наследуемости признаков, величины селекционного дифференциала, быстроты смены поколений, числа селективных признаков, так и от характера коррелятивных связей между ними.
Изучение коррелятивных связей между признаками пчелиных семей и выявление возможностей использования косвенного отбора в селекции пчел будут способствовать эффекту селекции.
Медопродуктивность является главным селективным признаком большинства селекционных программ, и поэтому изучение коррелятивных связей хозяйственно полезных и экстерьерных признаков непосредственно с медопродуктивностью представляет наибольший интерес.
Ниже приводятся данные, показывающие коррелятивные связи некоторых признаков с медопродуктивностью:

Зимостойкость пчелиных семей также положительно коррелирует с медопродуктивностью, хотя между оценками этих признаков проходит 3—4 месяца — практически весь активный пчеловодный сезон. Следовательно, по результатам зимовки еще ранней весной можно сделать предварительный прогноз медопродуктивности.
Значительной по силе положительной корреляцией связана с медопродуктивностью и среднесуточная яйценоскость маток, а также определяющиеся ею максимальная яйценоскость, общая масса пчел на период медосбора и летная деятельность пчел.
Из признаков, коррелирующих с медопродуктивностью, более подробного рассмотрения заслуживают такие сложные хозяйственно полезные признаки, как зимостойкость пчелиных семей и яйценоскость маток. В зависимости от целей селекции они могут быть и самостоятельными. Зимостойкость пчелиных семей складывается из целого комплекса более простых показателей, находящихся в коррелятивной зависимости (табл. 6).

Наполненность кишечника пчел экскрементами, положительно коррелирующая с такими показателями, как зимний отход пчел, расход корма за зиму, степень опоношенности гнезд, и отрицательно — с количеством печатного расплода весной, может служить в качестве сигнального признака успеха зимовки. Данные по этому признаку можно получать в результате взвешивания кишечников у пробных групп пчел, взятых еще в зимовнике; можно наблюдать и за динамикой накопления экскрементов в течение зимы
Степень опоношенности гнезд также можно использовать в качестве сигнального признака успеха зимовки, определяя ее как в зимовнике, так и сразу же после выставки при беглом весеннем осмотре пчелиных семей, когда погодные условия не позволяют производить полную разборку гнезд.
Количество печатного расплода при 1-м весеннем учете, отрицательно коррелируя с зимним отходом пчел, расходом корма за зиму, степенью опоношенности гнезд и наполненностью кишечника экскрементами, также может служить косвенным показателем успеха зимовки. Кроме того, этот показатель положительно коррелирует и с общей массой пчел, выращиваемых к медосбору (л=0,32 — 0,65). Было изучено влияние на показатели зимостойкости и некоторых других признаков (таб. 7).

Наблюдается тенденция к положительной связи этого признака со степенью опоношенности гнезд.
Активность каталазы ректальных желез пчел коррелирует отрицательно с каловой нагрузкой, расходом корма за зиму и имеет тенденцию к отрицательной связи со степенью опоношенности гнезд. Слабая положительная корреляция наблюдается между активностью каталазы ректальных желез пчел и количеством печатного расплода.
Увеличение процента сахарной добавки в зимних кормах приводит к снижению расхода корма за зиму, к уменьшению степени опоношенности гнезд и величины каловой нагрузки, и в то же время отрицательно сказывается на количестве печатного расплода весной. Отмечая положительное влияние замены части меда в зимних запасах сахаром на некоторые показатели зимостойкости, необходимо в то же время определить оптимальные размеры сахарной добавки применительно к определенным природно-климатическим условиям и конкретным породам пчел.
Зимостойкость пчелиных семей, выраженная комплексным баллом, оказывает определенное влияние непосредственно на медопродуктивность (см. табл. 7) и более выраженно влияет на яйценоскость пчелиных маток:

Несколько ниже значения коэффициентов корреляции зимостойкости с яйценоскостью (определяются за 36 дней до начала главного медосбора), и совсем незначительна корреляция зимостойкости с максимальной яйценоскостью непосредственно перед началом главного медосбора.
Максимальная яйценоскость связана сильной положительной корреляцией со среднесуточной яйценоскостыо (г=0,67 — 0,91), что дает основание судить о среднесуточной яйценоскости по значению максимальной. Это обстоятельство дает возможность при крупномасштабной селекции проводить экспресс-оценку общей яйценоскости по показателю максимальной. С максимальной яйценоскостью маток положительно коррелирует их масса (г=0,31), а с массой — длина и ширит 3-го стернита, т. е. размеры тела (г=0,56 — 0,77). Это дает возможность проводить косвенный отбор на яйце носкость по массе и размерам маток сразу же после их вывода.
Таким образом, изученные хозяйственно полезны признаки пчел представляют собой комплекс устойчивых взаимосвязей. Направление коррелятивных связей по годам не изменялось, а их сила колебалась незначительно. Анализ коррелятивных связей показывает возможность проведения косвенного отбора по селективным признакам и осуществление ранней оценки селенкционного материала.
Мы проанализировали также возможности косвенного отбора по качеству маток. Пчеловоды знают, что даже матки с известным происхождением и несомненно хорошей наследственностью не всегда показывают хорошую яйценоскость. Другими словами, ценные гены иногда могут реализоваться в посредственных и также плохих фенотипах. Причиной этому могут быть плохие условия содержания и кормления маток в семьях-воспитательницах, недостаток пчел-кормилиц, технические недоработки при прививке личинок и целый ряд других факторов, которые трудно учесть и устранить.
В этой связи возникает необходимость тщательного отбора получаемых маток по признакам, доступным для непосредственной оценки — внешнему виду, размерам, массе, т е. по фенотипу. Такой отбор повышает также результативность селекционной работы. Здесь вполне уместно известное изречение М.Ф.Иванова: «Хорошие генотипы следует искать среди хороших фенотипов».
Учитывая такую биологическую особенность пчелиных маток, как короткий срок жизни, особое значение приобретают методы ранней оценки маток по косвенным признакам — размерам маточников, массе неплодных маток по выходе из маточников и т. д.
Однако данные о корреляционных связях между экстерьерными и интерьерными признаками пчелиных маток, а также их яйценоскостью весьма противоречивы.
Так, П.М.Комаров (1937), К.Вейс (1972) не установили четкой связи между весом маток и числом яйцевых трубочек. Г.А.Аветисян (1967) обнаружил лишь слабую связь между этими признаками.
Л.А.Хидешели и Н.В.Мчедлишвили (1973), Ф.Таранов (1974) выявили у маток серых горных кавказских пчел, а Хорошилов (1973) у маток дальневосточных пчел высокую положительную корреляцию между весом маток и количеством яйцевых трубочек.
А.Н.Мельниченко и др (1968), А.С.Тришина и Н.Д.Шмелева (1974) провели подобную работу, рассматривая разные породы пчел, и в целом выявили слабую связь между весом маток и количеством яйцевых трубочек. Они нашли положительную связь между числом яйцевых трубочек и объемом маточников у кавказских, краинских и дальневосточных маток. Отмечена высокая положительная корреляция между числом яйцевых трубочек и глубиной маточников у итальянских и дальневосточных маток. В большинстве же случаев не наблюдается достоверной корреляции между числом яйцевых трубочек и размером маточников.
Меньше разногласий вызывает характер корреляции между объемом маточников и весом неплодных маток а также размерами тергитов и стернитов и весом маток. Здесь отмечается положительная корреляция. (Г.А.Аветисян и др., 1971; П.О.Губа, 1973; А.Л.Хидешели и Н.В.Мчедлишвили, 1973).
В результате наших исследований (Н.И.Кривцов, М.Д.Большакова) установлена положительная корреляция между массой маток и большинством экстерьерных признаков (табл. 8, 9). Особенно четкая взаимосвязь наблюдается между массой маток и размерами 3-го стернита.

Количество яйцевых трубочек более тесно связано с шириной 3-го тергита и 3-го стернита, с длиной 3-го тергита корреляции не обнаружено. Не удалось установить и связи массы неплодных маток с количеством яйцевых трубочек. В опыте маток взвешивали по выходе из маточников и каждую нумеровали. Нередко попадались матки, у которых в каждом яичнике насчитывали до 170 яйцевых трубочек, а весили они как 180 так и 210 мг. В то же время по 140 яйцевых трубочек в одном яичнике было, у маток массой 160, 180, 210, 220 мг.

Обнаружена положительная связь между глубиной маточников и массой неплодных маток, а также между объемом маточников и массой неплодных (г = 0,46) и плодных (г = 0,22) маток. Масса неплодных и плодных маток тесно коррелирует, что открывает возможности объективного раннего отбора по первому признаку.
В опыте 1985 г. масса 51 неплодной матки составила в среднем 206 мг, а масса этих же маток после того , как они стали плодными, достигла 229,5, т. е. увеличилась в среднем на 23,5 мг.
Хотя проследить влияние массы маток на их яйценоскость сложно из-за множества паратипических факторов, определяющихся внешними и внутренними условиями жизни пчелиной семьи, эти признаки положительно коррелируют. В наших исследованиях с максимальной яйценоскостью маток коррелировала их масса (взвешивали неплодных маток, г = 0,31 ±0,174, гс = 32). Отмечена незначительная корреляция массы неплодных маток с силой пчелиных семей на медосборе (г=0,41 rfc ±0,130, п—72; 1985 г.). Приведенные данные также свидетельствуют в пользу косвенного отбора на яйценоскость по массе неплодных маток. Однако если проанализировать влияние массы неплодных маток на максимальную яйценоскость плодных маток в узких пределах (от 195 до 225 мг), то здесь корреляция отсутствуем (г = —0,11, гс = 40; 1985 г.).
На наш взгляд, можно выделить два основных признака, предварительно характеризующих качество маток,— их массу и количество трубочек в яичниках. С учетом изученных корреляций можно составить следующую схему косвенного отбора качественных маток, отбраковка неполноценных маточников; отбор неплодных маток по оптимальной массе (220 мг) и размерам 3-го тергита и стернита, особенно по их ширине (в производственных условиях отбор проводится визуально по размерам тела); контроль за наличием оставшегося корма в маточниках по выходе матки (отсутствие корма показывает неполноценность кормления); контроль за внешним видом маток.
Основная и окончательная оценка качества маток проводится непосредственно по их яйценоскости в пчелиных семьях.
В то же время, следует иметь в виду, что масса плодных маток в пределах, близких к стандарту, может и не коррелировать с их максимальной яйценоскостью и силой пчелиных семей на медосборе (в 1985 г. мы выявили даже тенденцию к отрицательной корреляции между этими признаками). Это дает основание предполагать, что более плодовиты матки с массой, приближающейся к 220 мг (стандарт), а не к 250 мг (в опыте). Следовательно, целесообразно отбирать маток оптимальной массы, бракуя легких (ниже стандарта) и не стремясь к отбору чересчур тяжелых (больше 250 мг).
Таким образом, значения генетико-популяционных параметров основных селективных признаков среднерусских пчел, таких, как фенотипическая и генотипическая изменчивость, наследуемость, повторяемость, дают основание рассчитывать на определенный успех селекции. Их необходимо анализировать на всех стадиях селекционного процесса.
Также должен сопутствовать селекционному процессу анализ корреляций, что позволяет более объективно оценивать селекционные признаки, дает возможность, прогнозирования развития одних из них по другим, а также осуществления косвенного отбора.
ОРГАНИЗАЦИЯ, МЕТОДЫ И ПРОГРАММЫ СЕЛЕКЦИИ ПЧЕЛ
Методы селекционной работы
Медоносные пчелы — насекомые с высоким уровнем общественной жизни, живущие семьями, состоящими из одной самки (матки), нескольких десятков тысяч ее дочерей (рабочих пчел) и нескольких сотен сыновей (трутней), которые появляются в благополучных семьях только летом и выполняют функцию размножения (См. Биология пчелиной семьи. ).
Трутни гаплоидны, т. е. они развиваются из неоплодотворенных яиц, тогда как матки и рабочие пчелы диплоидны — они происходят из яиц, оплодотворенных спермой, хранящейся в сперматеке матки и полученной от нескольких (до 15) трутней. Спаривание маток и трутней происходит в воздухе, иногда на значительном расстоянии от пасеки, что затрудняет его контроль (См. Процесс спаривания. ).
В то время как трутень спаривается один раз в жизни и только с одной маткой, матка чрезвычайно плодовита и может откладывать 2—3 тыс. яиц в сутки. Образно говоря, трутни представляют собой гаметы от матки. Отмеченные особенности показывают целесообразность селекции по женской линии.
Многие хозяйственно полезные признаки медоносной пчелы (медопродуктивность, воскопродуктивность) являются результатом деятельности рабочих пчел семьи, и селективная ценность этих признаков оценивается не через участвующих в спаривании маток и трутней, а через рабочих пчел. Таким образом, пчелиная семья в целом представляет собой своеобразный и оригинальный селекционный объект.
Как и в других отраслях животноводства (конечно, с учетом специфических особенностей), в пчеловодстве используют методы чистопородного разведения и скрещивания.
Подчеркивая творческое значение селекционной работы вообще, известный советский генетик Н.И.Вавилов (1934) писал: «Селекция по существу есть вмешательство человека в формообразование животных и растений; другими словами, селекция представляет собой эволюцию, направляемую волей человека». Формы и методы селекционной работы могут быть различны: одни селекционеры предпочитают селекцию с использованием нескольких пород, другие являются сторонниками чистопородного разведения (См. Старая история о наилучшей пчеле...).
Еще известный швейцарский ученый-пчеловод Крамер (1898—1899), рассуждая о выборе породы, отмечал, что местная пчела, «...которая в течение целых тысячелетий приноравливалась к условиям нашего климата и акклиматизированная подходящими способами, должна уже по одному тому представлять из себя самый благородный продукт для разведения, так как она менее всего склонна к вырождению».
Рассмотрим схемы селекционной работы в некоторых странах с развитым пчеловодством.
Задача племенной работы в пчеловодстве — постоянное улучшение пород пчел (См. Породы пчел.). Следствием такого улучшения является развитие основных хозяйственно полезных признаков (медопродуктивности, воскопродуктивности) и их устойчивая передача из поколения в поколение. Хотя показатели продуктивности пчел складываются из множества факторов (длина хоботка рабочих пчел, нагрузка их медового зобика, продолжительность жизни, работоспособность, предприимчивость в отыскании источников медосбора, устойчивость к заболеваниям, зимостойкость, плодовитость маток и др.) и зависят от них, все же селекцию целесообразно вести не по этим отдельным показателям, а по результатирующему признаку, в частности медопродуктивности.
Чтобы оценка и отбор пчелиных семей были объективными, необходимо их проводить среди возможно большего количества сравниваемых пчелиных семей.
В любом случае первым этапом селекционной работы должно быть получение линий на основе чистопородного разведения, максимальное использование их генетических особенностей для достижения наибольшего хозяйственно полезного эффекта. Пути и формы такой работы могут быть различны. Однако развитие таких линий ограничено, и наступает этап, когда возникает необходимость проведения межлинейного скрещиваиия как внутри породы, так и между породами
F.Kobel (1968), следуя концепции Крамера о преимуществах чистопородного разведения местных пчел, тщательно переработал девятое издание его книги, изменив название на «Чистопородное разведение медоносной пчелы». Это фундаментальный труд, полезный не только в качестве практического руководства по чистопородной селекции для швейцарских пчеловодов, но интересный для всех селекционеров.
Важнейшим моментом Ф.Кобел считает составление тщательного плана селекционной работы. Решающее значение имеет отбор племенных пчелиных семей. При этом следует контролировать также продуктивность семей с матками-сестрами,. тетками и др., т. е. вести отбор по происхождению и по качеству потомства. Средством для проведения такого отбора является балльная оценка. После оценки исходного материала необходимо выбрать не менее шести племенных маток, вывести от них дочерей и сформировать группы семейств. После двухлетнего испытания отбирают лучших.
При разведении по линиям вследствие инбридинга может ослабляться жизнеспособность пчелиных семей. В таком случае встает вопрос об «освежении крови». Хорошая селекционная работа невозможна без использования изолированных случных пунктов. Однако замена маток на соседних пасеках и без надежной изоляции (См. Гаранты чистопородности при репродукции в неизолир...), по мнению Ф.Кобела, может обеспечить в течение нескольких поколений хороший селекционный эффект.
К.Dreher (1976) дал обстоятельную и обобщенную характеристику методам чистопородного разведения и скрещивания в пчеловодстве. Среди предпосылок успешной селекционной работы он выделяет ясность селекционной цели, наивысшую продуктивность при возможно более низких затратах, единообразие состава и надежность наследования ценных признаков. Селекционер может влиять на генотип в результате отбора (накопление плюс-вариантов и исключение минус-вариантов) или используя эффект гетерозиса (См. О гетерозисе и вреде не контролируемой гибридизаци...). При этом следует иметь в виду, что полученные в последнем случае изменения не наследственны. Различные методы внутрипородной селекции можно классифицировать следующим образом.
Инбридинг (инцухт) предполагает: тесное линейное разведение, приводящее через несколько поколений к образованию инбредных линий; широкое линейное разведение; тесное групповое разведение (например, внутри штаммов Тройзен, Скленар); широкое групповое разведение (например, внутри экотипов альпийской карники).
Аутбридинг (аусцухт) предполагает; планомерное 4-линейное скрещивание инбредных линий; планомерное скрещивание различных по происхождению групп одного экотипа; планомерное скрещивание различных экотипов.
Среди методов межпородного скрещивания можно выделить получение и промышленное использование F-1,многократное пользовательное скрещивание, ротационную селекцию, вытеснительное скрещивание, породообразующее скрещивание.
Одним из первых исследование тесного инбридинга у пчел провел современник Дзержона и Берлепша Ф.Фогель, который от двух семей египетских пчел вывел множество поколений. Начиная с 6—7-го поколения пчелиного расплода становилось все меньше, семьи делались нежизнеспособными.
После второй мировой войны Г.Виддель из штамма Скленар путем близкородственного спаривания вывел свой штамм Альба. Он достиг крайнего усиления признаков карники, но при этом также значительно снизилась жизнеспособность пчел. Такая же проблема возникла у доктора Кейла из США, получившего для фирмы «Дадан и сыновья» несколько инбредных линий. Содержание таких линий для последующего кроссирования было сопряжено с большими издержками.
Как только вследствие инбридинга происходит обеднение генов, повышается (до 50%) выпадение расплода пчел. Правда, как уже было сказано выше, против нежелательных последствий инбридинга у медоносных пчел выработался целый ряд приспособлений: спаривание половых партнеров происходит далеко от своей пасеки и в основном с неродственными партнерами; молодые матки за 1—3 брачных вылета спариваются в среднем с 8—9 трутнями; гаплоидность трутней способствует тому, что вредоносные гены у них не могут оставаться в скрытом состоянии, они проявляются внешне (белоглазость, бесфасеточность глаз). Такие трутни не выдерживают конкуренции в местах их сбора.
Нетесный инбридинг при широком линейном разведении (спаривание кузинов и родственников с более отдаленным родством) вполне приемлем для селекций при наличии надежных пунктов спаривания или инструментального осеменения.
Тесное групповое спаривание осуществляется внутри определенной популяции. Пока штаммы не сужаются по линиям, опасности инбридинга нет.
Успешно можно практиковать широкое групповое разведение.
Продукты скрещивания инбредных линий отличаются более высокой продуктивностью за счет эффекта гетерозиса. Однако этот эффект неустойчив, и через несколько поколений продуктивность снижается до уровня исходного материала и ниже. Широко известны гибриды кукурузы, полученные на основе инбредных линий. По этому же принципу были созданы гибридные штаммы пчел Старлайн и Миднайт в США.
Достаточно распространено и планомерное скрещивание внутри экотипа, а также различных экотипов внутри породы. Этот метод также дает возможность получать значительный эффект гетерозиса, но так бывает далеко не всегда, и поэтому в каждом конкретном случае надо проводить специальные исследования.
С середины прошлого столетия начался завоз иноземных пород пчел, что повсеместно привело к бесплановой метизации и уничтожению генофонда местных пород См. Экологический гетерозис пчелиных семей и ошибки по...). Содержание чистых пород и плановое получение помесей первого поколения требует предварительных расходов и четкой организации (См. О гетерозисе и вреде не контролируемой гибридизаци...).
В пчеловодстве возможны и многократные межпородные скрещивания. Этот способ успешно применил Адам при создании бакфастовской пчелы. При этом пчелы итальянской породы скрещивались с местными английскими пчелами.
Следует отметить также вытеснительное скрещивание, довольно широко используемое в пчеловодстве. При этом способе в целом хорошая порода, имеющая лишь некоторые недостатки, скрещивается с менее ценной, но не имеющей этих недостатков. Последующие генерации скрещивают снова с предпочитаемой породой. Стихийное вытеснительное скрещивание происходит при скрещивании местных пчел с завозными породами.
Хочется предостеречь в селекционной работе от необдуманного скрещивания, метизации ценных популяций местных пчел, наилучшим образом приспособленных к условиям среды. Исходя из учета одного лишь показателя — медопродуктивности, /. М. Cornuet (1981) предложил следующий упрощенный план ее повышения.
В каждом поколении отбирают 10 пчелиных семей. От каждой из этих семей получают маток для организации 12 семей-дочерей. Таким образом получают 120 пчелиных семей от 10 основательниц. Эти 120 семей распределяют на четыре точка по 30 семей. По результатам оценки медопродуктивности этих 120 семей отбирают из них 10 для дальнейшей селекции. Отбор основывается как на собственной ценности семьи, так и на средней ценности семей данной группы. Спаривание естественное, но принимаются меры для выращивания большого количества трутней нужного происхождения. Схема данной селекции строится на основе местной популяции с обязательным мечением маток.
J.P.Chaplean (1984), рассматривая различные подходы к селекции пчел, отметил, что их медопродуктивность нельзя повысить, не проводя отбора по качеству потомства. При этом можно подбирать сотни пчелиных семей по длинному списку критериев, и можно ограничиться лишь несколькими признаками и небольшим количеством пчелиных семей (в зависимости от возможностей селекционера). Поскольку контроль зa генотипом трутней осуществить трудно, да и спариваться они могут раз в жизни, приходится, как говорилось выше, отдавать предпочтение селекции по женской линии. Однако и в этом случае можно значительно улучшить качество потомства. Такая селекция дает эффект уже у следующего поколения трутней.
Этот автор предлагает два метода селекции — групповую, когда от одной или нескольких отобранных маток получаются все остальные (и так из поколения в поколение) при естественном спаривании, и групповую с группами. В последнем случае матки в каждой подгруппе — сестры. Ежегодно новая матка-родоначальница выбирается внутри каждой подгруппы и ее дочери продолжают линию. Это, собственно, схемы массового (фенотипического) и генотипического отбора.
J.Vaillqnt (1982) вслед за другими авторами призывает учитывать наследственность трутней и считает, что она имеет не меньшее значение, чем наследственность маток в формировании генотипа женских особей потомства. Это не отрицают и другие селекционеры, однако подобное возможно при наличии надежных изолированных случных пунктов или при использовании инструментального осеменения.
К тому же, занимаясь селекцией трутней, следует иметь в виду, что у них могут быть лишь дочери, деды и внуки. Матки чистой породы производят генетически идентичных трутней той же породы. Если в определенную местность завозят матку другой породы, она дает трутней своей породы, которые будут спариваться с местными неплодными матками, вытесняя прежнюю породу. В одной пчелиной семье самки, происходящие от одного трутня, имеют коэффициент родства 3/8 вместо ]/4 при наличии двух диплоидных родителей, поэтому Лейдлоу предложил называть их суперсестрами. Риск близкородственного скрещивания компенсируется многократным спариванием маток. В противном случае близкородственное скрещивание привело бы к снижению резистентности рабочих пчел к заболеваниям, уменьшению активности их летной деятельности, сокращению продолжительности жизни трутней, появлению генетически «пестрого» расплода. Чтобы этого не случилось, необходимо оперировать несколькими отобранными линиями.
Признаки отбора пчелиных семей для вывода трутней соответствуют таковым при выводе маток.
Пчелиная семья в период максимального развития отстраивает до 14% трутневых ячеек, а в конечном итоге их бывает 2—3% от общего количества рабочих пчел. Если нет надежного изолированного пункта спаривания, необходимо применять следующие приемы: стимуляцию селекционных трутней к вылету в определенное время, когда другие трутни не летают, спаривание вне сезона, создание мощного трутневого барьера за счет преимущественного вывода нужных трутней.
W.G.Rothenbuhler (1980) считает, что любая программа улучшения породы пчел должна состоять из трех этапов: полевых испытаний, селекционной работы и коммерческого производства улучшенных пород пчел. При этом полевые испытания должны проводиться именно в тех районах, где порода будет использоваться. Задача селекционера — спланировать порядок полевых опытов, отобрать пчелиные семьи для селекции, решить, когда использовать инструментальное осеменение, а также как поддерживать и использовать улучшенную породу. По мнению этого автора, успех осуществления программы правильного определения района испытания пчелиных семей зависит от признаков отбора (как правило, основной — медопродуктивность), количества пчелиных семей, испытываемых в каждом поколении (в среднем 50); а также процента из них, необходимого для дальнейшей селекции (автор,советует брать 10%). Также важны организация полевых испытаний оплодотворенных естественным путем маток, продолжение подобной работы в следующем поколении, сравнение новой линии с используемыми на пасеке пчелами, размножение и реализация маток этой линии в случае ее превосходства.
Издавна между селекционерами-пчеловодами идет дискуссия о том, использовать ли чистые породы или межпородные гибриды.
W.С.Roberts, О.Mackensen (1951) показали, что противоречий в этих методах нет. Идеальная система скрещивания — это селекция и поддержание чистых, но неродственных линий и их скрещивание для получения улучшенных гибридов. Выводимые инбредные линии слабее неинбредных, однако их правильные комбинации, как уже говорилось, дают эффект гетерозиса.
Н.Cale, J.Н.Gowen (1956) проанализировали генетический материал из четырех инбредных линий пчел. Ими получены различные результаты, позволяющие сделать вывод о том, что не все гибриды одинаково ценны, и приходится изучать и учитывать комбинационную способность инбредных линий.
При хорошей организации и планировании чистопородного разведения удается получать значительный селекционный эффект без риска метизации и потери ценных породных качеств. Чистопородная селекция основывается на долгосрочных, рассчитанных на несколько лет, комплексных программах.
Селекционные программы, используемые в странах с развитым пчеловодством
Знакомство с программами хотелось бы начать с опыта селекционеров Австрии. Систему племенной работы в этой стране регламентирует Австрийский союз пчеловодов (Н.И.Кривцов, Г.Н.Котова, 1983). Основным принципом его деятельности является тесная связь науки и практики — Института пчеловодства как центра племенной работы и пчеловодов-промышленников на местах. В этом комплексе ежегодно задействовано 1500—2000 пчелиных семей. В Австрии разводят в основном аборигенных краинских пчел, а в северных районах — темных лесных. Племенных маток передают на частные пасеки для испытания. От каждой проверяемой по качеству потомства матки оценивают по 5—15 маток-дочерей. Австрийские селекционеры считают, что лучше сократить число дочерей в группах, но проверить по качеству потомства как можно больше маток-рекордисток. Оценку ведут 1—2 года. Важнейшим селективным признаком при этом является медопродуктивность. Ее учитывают пчеловоды-практики на своих пасеках и по специальной форме передают результаты в институт для анализа. Если медопродуктивность устраивает селекционеров, то приступают к оценке других признаков, и прежде всего чистопородности. Чистопородность по экстерьерным признакам устанавливает институт.
Таким образом, на частных пасеках, расположенных в различных условиях, проводится конкурсное испытание племенного материала, получаемого из института. Данные сравнивают с показателями местных линий. Если в результате оценки на пасеках-испытательницах выявляется ценный для селекции материал, то его передают институту в виде яиц и молодых личинок для последующей репродукции.
Опираясь на сеть пасек-испытательниц, институт ведет 7 линий по 100 пчелиных семей. Спаривание маток институт осуществляет на станциях спаривания, расположенных в изолированных долинах Альп. Рассылаемых маток метят метками различной формы и цвета, обозначающими их происхождение и год рождения. Постоянная пропаганда и реклама результатов селекции способствуют повышенному спросу на племенных маток. Пчеловодам, которые аккуратно и добросовестно присылают сведения о результатах испытания, поставляют маток на льготных условиях.
В ФРГ (Баварский институт пчеловодства) селекционируют линии (штаммы) краинских пчел, которые оценивают на трех испытательных пунктах (К.Weiss, 1982). На каждом пункте испытывают 10 групп по 10 пчелиных семей в каждой. Испытания, не считая год подготовки, когда подсаживают маток, продолжаются 2 года.
Продуктивность оценивают и регистрируют по каждой пчелиной семье отдельно. Детальные отчеты даются за каждый год испытания, а после окончания испытания институт представляет в специальные издания по пчеловодству обобщенный отчет.
В бывшей ГДР селекционеры использовали карнику (G.Pritsch, 1983). Были созданы многочисленные случные пункты с отобранным трутневым составом определенного происхождения. С мест испытания сюда возвращали оцененный племенной материал. Работу с каждой линией обеспечивали 5—6 пунктов, образующих комплекс. Каждый такой комплекс представлял собой замкнутую селекционную систему, что давало возможность исключить бесплановые скрещивания. Всего же действовало 60 случных пунктов, объединенных в 12 замкнутых комплексов. Племенные пчелиные семьи отбирались по принципу чистопородности в соответствии со стандартом. В радиусе 10 км от случного пункта не должно было быть инородного материала
Именно на основе комплексов случных пунктов и строилась работа по планам ступенчатого линейного разведения (линии оценивали в нескольких, не менее чем в трех, поколениях). Здесь же получали необходимое количество чистопородных племенных маток, контролировали кочевки в зонах чистопородного разведения и оценивали чистопородность племенного материала.
В Чехии (V.Vesely, 1982) селекционную работу координирует Институт пчеловодства в Доле. Непосредственное участие в организации этой работы принимает Чешский союз пчеловодов. Все пасеки делятся на следующие категории: областные, признанные, разведенческие, зарегистрированные, товарные.
Областные пасеки относятся к высшей категории. Здесь селекционные достижения внедряются в практику, количество пчелиных семей на таких пасеках должно быть не менее 40.
На признанных пасеках (размером также не менее 40 пчелиных семей) обеспечивается контроль над спариванием маток. Селекцию здесь ведут методом индивидуального отбора. Пчеловоду необходимо доказать свою профессиональную пригодность к работе на признанной пасеке. Это значит, он должен не менее чем в трех поколениях пчел проводить оценку признаков, в том числе и экстерьерных, на соответствие стандарту, обеспечивать контроль над спариванием, а также подтвердить, что в течение двух лет средний показатель медопродуктивности составлял не менее 80% от показателя областной пасеки. Признание его профессиональности действует 10 лет, затем проводят повторную проверку по перечисленным выше критериям.
Разведенческие пасеки размножают племенной материал какой-либо признанной или областной пасеки. Здесь ведут массовый отбор трутней, ежегодно заменяют не менее 60% маток. Такие пасеки также насчитывают не менее 40 пчелиных семей при непременном условии ежегодной продажи не менее 100 маток на сторону. Спаривание маток проводят на территории пасеки. Заявитель на статус разведенческой пасеки представляет карту местности с указанием местонахождения своей пасеки и пасек других пчеловодов в радиусе 5 км, которые будут брать маток именно у него и заменять ими ежегодно не менее 20% маток на своих пасеках.
Зарегистрированные пасеки насчитывают не менее 20 пчелиных семей, на них проводят массовый отбор.
Товарные пасеки могут выращивать маток только для собственных нужд.
В институте и на областных пасеках организована специальная служба, обеспечивающая спаривание привезенных в нуклеусах неплодных маток.
В Румынии (М.Щербан, М.Драган, 1984) по результатам оценки на конкурсах, по данным научных сотрудников, пчеловодов на высокопродуктивных пасеках, ежегодно выявляют маток — рекордисток по медопродуктивности. Оценку пчелиных семей проводят на испытательных и селекционных пасеках, организованных на зональном уровне. Кроме основного признака, медопродуктивности, учитывают яйценоскость, миролюбие, поведение на сотах, ройливость и др. Отличившиеся по комплексу признаков пчелиные семьи передают на элитные пасеки или в центры, предназначенные для создания и совершенствования новых линий или кроссов. Работа на этом уровне относится непосредственно к компетенции института. Элитные пасеки и центры производят материал высокой племенной ценности, они постоянно обмениваются ценными, по собственным результатам и результатам потомства, матками с другими пасеками страны и пасеками-испытательницами.
Линии со стабильными признаками и лучшие кроссы представляют специальной комиссии для официального признания и включения в книгу республиканского центра селекции.
Представление о системе селекционной работы в США можно получить, познакомившись с работой фирмы «Дадан и сыновья» и генетико-селекционной лаборатории в Батон-Руже (П.Уйтрелл, 1976; А.Харнаж, Т.Росс, Т.Дадан, Г.К.Кейл, 1976; G.Claerr, 1977).
Программа селекции в США предусматривает получение и сохранение инбредных линий и широкое использование в производстве межлинейных кроссов. Для получения 2-, 3-, 4-линейных кроссов используют маток с высокой степенью родства.
Получение планомерных гибридов стало возможным в США с развитием техники инструментального осеменения пчелиных маток (F.Е.Moeller, 1976). Для того чтобы матки также были гибридами, проводили многократные (3—6) межпородные скрещивания. Инбредные линии были произведены от нескольких пород, затем их комбинации испытывались на продуктивность. Всей программе селекции сопутствовал непрерывный отбор по медопродуктивности, устойчивости к американскому гнильцу и некоторым другим характеристикам (злобливость, ройливость, склонность к прополисованию, качество расплода, характер печатки меда, окраска тела).
Чтобы сохранять преимущества гибридов, необходимо поддерживать в чистоте инбредные линии и всякий раз получать гибрид первого поколения. Поддерживать же инбредные линии, имеющие низкую жизнеспособность, непростая задача. Поэтому следует избегать высокой степени инбридинга исходных пород.
Маток, дающих пчел межлинейных кроссов, распределили между участвующими в программе пчеловодами-селекционерами, которые их испытали и отобрали племенной материал для последующих межлинейных скрещиваний. Чистота кроссов гарантировалась инструментальным осеменением.
Для выполнения комплексных селекционных программ считается целесообразным иметь специальную пасеку с лабораторией сохранения и поддержания инбредных линий, пасеку для получения межлинейных кроссов первого поколения и сеть пасек для оценки их селекционной ценности. Так, широко распространенный в США гибрид Старлайн основывается на четырех инбредных линиях итальянских пчел. Ежегодно выводят новые линии, которые сравнивают с уже существующими. Оказавшиеся лучшими в результате оценки линии становятся источником получения маток и трутней для очередного поколения. Этот племенной материал пчеловоды-селекционеры возвращают фирме для получения коммерческих кроссов. Такая кооперация фирмы с пчеловодами-селекционерами очень эффективна.
Обобщая зарубежный опыт, следует отметить рациональные стороны чистопородной селекции.
Работу репродукторов, как правило, курируют научные учреждения селекционного профиля, что позволяет размножать племенной материал, постоянно улучшаемый в процессе селекции.
Существует постоянная взаимосвязь между центрами селекции и репродукторами, с одной стороны, и заказчиками маток — с другой. Это дает возможность анализировать результаты, получаемые на местах, и, таким образом, отбирать наиболее ценный материал для дальнейшей селекции и репродукции. Благодаря организации обратной связи небольшой коллектив селекционеров может решать задачи крупномасштабной селекции.
Селекционная работа проводится на основе относительно крупных массивов пчел одного происхождения, что исключает возможность бесплановой метизации. В репродукторах чистота племенного материала поддерживается периодическим инструментальным осеменением, если нет надежных изолированных случных пунктов.
В основе селекции — четкие программы, регламентированные соответствующими органами.
Методы селекционной работы в России
В нашей стране чистопородная селекция строится либо на основе массового, фенотипического отбора, либо с использованием индивидуального, генотипического отбора с проверкой маток по качеству потомства и выведением линий.
Массовый отбор является наиболее доступным и испытанным способом улучшения хозяйственных и племенных качеств пчелиных семей. Его сущность заключается в планомерной выбраковке худших пчелиных семей и интенсивном размножении лучших. Как уже говорилось, отбор ведется по медопродуктивности, однако учитываются и такие хозяйственно полезные признаки, как зимостойкость, устойчивость к заболеваниям, плодовитость маток и сила пчелиных семей, ройливость, воскопродуктивность и т. д.
В конце сезона все пчелиные семьи оценивают по перечисленным признакам и распределяют на 3 группы. В первую группу выделяют 10—15% лучших пчелиных семей, в 1,5—2 раза превосходящих средние показатели пасеки по медопродуктивности, хорошо перезимовавших, без признаков заболеваний, с высоколодовитыми матками и т. д. Кроме того, семьи пчел, выделяемые в первую (племенную) группу, должны быть чистопородными, т. е. отвечать по всем своим признакам типу дайной породы. Так, у среднерусских пчел окраска тела не должна иметь желтизны, печатка меда должна быть белой («сухой») и т д.
Ко второй группе, самой многочисленной, относят пчелиные семьи со средними показателями. К третьей группе (10—15% от общего числа пчелиных семей) относят худших по остальным показателям и наименее продуктивных. Семьи третьей группы необходимо выбраковывать сразу же, т. е. в конце сезона, или в крайнем случае в следующий сезон.
В следующий сезон в качестве племенных используют семьи первой группы. В семьях второй группы сменяют маток на маток, выведенных от семей первой группы. Для смены старых маток в семьях первой группы и для подсадки во вновь формируемые отводки используют также только маток, выведенных от лучших семей первой группы.
В выделенных из первой группы отцовских семьях создают наилучшие условия для выращивания трутней— подставляют 1—2 трутневых сота, тщательно утепляют гнезда, систематически дают стимулирующую подкормку. Одновременно с этим применяют все меры, чтобы не допустить вывода трутней во всех остальных семьях пасеки.
Улучшение породы будет более успешным, если такую же племенную работу ведут и на окружающих пасеках, расположенных в зоне с радиусом до 12—15 км. В противном случае возможно нежелательное спаривание значительной части маток с трутнями из малопродуктивных семей соседних пасек.
Целесообразно выводить молодых маток от пчелиных семей, неродственных отцовским, а также через 1 - 4 года производить обмен племенными пчелиными семьями между пасеками, разводящими одну и ту же породу пчел и находящимися друг от друга не ближе, чем в 25—30 км, чтобы избежать родственного спаривания маток и трутней.
Массовый отбор, проводящийся систематически из поколения в поколение, улучшает также племенные качества пчелиных семей, т. е способность устойчиво передавать своим дочерним семьям высокую продуктивность и другие ценные хозяйственно полезные признаки.
Описанная схема применима, разумеется, к пчелам всех пород. Неоднократно отмечалось, что массовый отбор на пасеках среднерусских пчел приводил к значительному повышению их медопродуктивности. Так, Н.Солодкова (1951), Т.С.Жданова (1970) сообщали, что в результате массового отбора среднерусские пчелы татарской популяции при благоприятных условиях климата и взятка могут собирать за сезон до 100 кг меда на каждую пчелиную семью.
Индивидуальный генотипический отбор — более высокая форма племенной работы. Однако исходные позиции остаются такими же, как и при применении массового отбора. Индивидуальную селекцию ведут, как правило, в научно-исследовательских учреждениях по пчеловодству, располагающих необходимой для этого базой и силами.
Имеются примеры чистопородной селекционной работы со среднерусскими пчелами. Так, Н.Н.Гранкин (1978), занимающийся селекцией среднерусских пчел орловской местной популяции, выделил линии № 39 и 7, превосходящие контроль по медовой продуктивности соответственно на 28,5 и 39,4%, а по максимальной яйценоскости — на 39,4 и 32,5%. Этим селекционером апробированы и некоторые варианты внутрипородных межлинейных гибридов среднерусских пчел, также оказавшихся эффективными.
И.В.Шафиков (1978) проводил похожую работу с аборигенной бурзянской бортевой пчелой. В результате селекций им была выделена линия № 79 с повышенной на 25—35% плодовитостью маток, медовой и восковой продуктивностью пчелиных семей. В абсолютном выражении в проведенных испытаниях пчелиные семьи с матками-дочерьми, родоначальницы линии № 79, собрали меда на 8,3 кг больше, чем семьи контрольной группы. Пчелиные семьи с матками-внучками линии № 79 превышали семьи контрольной группы на 9 кг.
В отличие от других отраслей животноводства, где работа ведется, как правило, с мужскими линиями, в пчеловодстве основное внимание уделяется маткам и, соответственно, женским линиям. Это вызвано такими биологическими особенностями медоносных пчел, как большая плодовитость маток (до 2000 и более яиц в сутки), их скороспелость (от племенной матки за летний сезон можно получить и дочерей и внучек). Принимается во внимание и то, что трутни способны к спариванию лишь один раз.
Начало работы по проверке качества потомства пчелиных маток было положено исследованиями пчело-генетической лаборатории сектора генетики и селекции Всесоюзного института животноводства (ВИЖ) в 1930—1935 гг. под руководством академика А.С.Серебровского. После значительного перерыва селекционные работы были возобновлены, и в 70-е гг. был успешно применен метод проверки маток по качеству потомства при селекции дальневосточных, серых горных кавказских, карпатских пчел (Г.А.Аветисян, В.А.Губин, И.К.Давыденко, 1969; Ю.И.Макаров, 1969; В П.Кидалов, 1971, и др.). Все селекционеры отмечали большие возможности линейного разведения пчел различных пород с оценкой маток по качеству потомства, однако исследователи в большинстве случаев ограничиваются получением и испытанием одного-двух поколений.
СЕЛЕКЦИЯ СРЕДНЕРУССКИХ ПЧЕЛ
Как уже было отмечено, исходным материалом для селекции пчел среднерусской породы послужили 92 пчелиные семьи татарской популяции, завезенные из Сабинского района Татарии. Кстати, эта популяция в настоящее время успешно разводится в Мамадышском и Сабинском районах Татарии, где организован заказник среднерусских пчел.
В селекционном процессе участвовали только чистопородные пчелиные семьи (табл. 10). Племенное ядро линии № 44 было размещено в условиях относительной пространственной изоляции (радиусом около 3 км), и поэтому для сохранения чистопородности использовались дополнительно такие приемы, как изоляция во времени, создание мощного трутневого барьера, жесткий отбор по окраске хитина и экстерьерным признакам пчел. В результате этого был сформирован относительно устойчивый чистопородный массив среднерусских пчел в районе опытного поля.
 Ниже приводятся значения наиболее характерных экстерьерных признаков пчел исходной популяции по сравнению с усредненными данными восьмого и девятого поколений линии № 44 (осенняя популяция), полученными на 10 производственных пасеках массива (табл. 11).
Ниже приводятся значения наиболее характерных экстерьерных признаков пчел исходной популяции по сравнению с усредненными данными восьмого и девятого поколений линии № 44 (осенняя популяция), полученными на 10 производственных пасеках массива (табл. 11).Как видно из табл. 11, за 19 лет селекционной работы изменения экстерьерных признаков не произошло, они остались в рамках стандарта породы. Отмечено некоторое увеличение их фенотипической изменчивости (по значениям Cv%), однако значительную долю этого увеличения можно отнести за счет издержек при расчетах. Многолетний опыт свидетельствует о том, что даже в условиях недостаточной пространственной изоляции и при большой насыщенности пчелиных семей можно успешно сохранять и размножать чистопородный материал.

Основываясь на сделанных ранее расчетах, основным селективным признаком выбрали медопродуктивность. Поскольку зимостойкость связана с медопродуктивностью положительной корреляцией, она достаточно высока у среднерусских пчел, и задача состоит лишь в контроле за ее уровнем, а не в селекционном улучшении этого признака. К тому же отбор на медопродуктивность косвенно способствует отбору на зимостойкость.
Яйценоскость пчелиных маток (как среднесуточная, так и максимальная) также связана с медопродуктивностью положительной корреляцией (г=0,39 — 0,91), но этот признак может считаться, в зависимости от цели, и самостоятельным селективным (например, в хозяйствах разведенческого направления).
На первых этапах селекции отбор на медопродуктивность был очень жестким: на племя оставляли пчелиные семьи, превосходящие показатели сверстниц на 2q. В дальнейшем, по мере того как исчерпывалось генотипическое разнообразие, отбирали просто лучшие по медопродуктивности пчелиные семьи. В каждом поколении для репродукции последующего поколения отбирали 3—5 маток-рекордисток.
Селекционная работа проводилась на фоне относительно скудного главного медосбора с гречихи и сорно-полевого разнотравья, свойственного для центральных областей Нечерноземья, однако схема опыта дала возможность сделать объективные выводы об относительной ценности испытываемых по качеству потомства пчелиных семей аутбредной по трутням линии № 44.
В табл. 12 приведены данные о медопродуктивности племенных пчелиных семей этой линии.
 Для репродукции очередного поколения линии, за исключением первых двух, отбирали 3—5 пчелиных семей. Показатели медопродуктивности различны по годам, так как сильно зависят от медосборных условий сезона.
Для репродукции очередного поколения линии, за исключением первых двух, отбирали 3—5 пчелиных семей. Показатели медопродуктивности различны по годам, так как сильно зависят от медосборных условий сезона.При планировании опыта по Е.К.Меркурьевой (1970) определили численность дочернего потомства для оценки маток по медопродуктивности их дочерей. При q признака равной 5 кг и точности А в размере 3 кг, при критерии достоверности t 0,95=1,96 она равна 21 пчелиной семье:

Обращает на себя внимание тот факт, что для репродукции F-6 и F-7 были взяты пчелиные семьи, почти не отличающиеся от среднелинейных показателей: вариабельность признака к F-6—F-7 сильно снизилась в результате отбора. В предыдущих поколениях удавалось отобрать пчелиные семьи, в 1,5—2 раза превосходящие среднелинейные показатели. В табл. 13 приведены данные о медопродуктивности девяти поколений линии, полученных от родоначальниц, охарактеризованных в табл. 12.

Показатели медопродуктивности невысоки, однако селекционный прогресс очевиден. Во всех поколениях отмечается превосходство как над сверстницами, так и над исходной популяцией.
Яйценоскость пчелиных маток в решающей мере определяет жизнедеятельность пчелиной семьи в целом. Этот важнейший хозяйственно полезный признак может рассматриваться, как говорилось выше, и как самостоятельный селективный, имеющий определяющее значение при разведенческой специализации пчеловодства, и как косвенный — при селекции на другие признаки, в частности на медопродуктивность.
У пчел среднерусской породы в условиях среднелетнего главного медосбора связь между среднесуточной яйценоскостью за 36 дней до начала главного медосбора и медопродуктивностью проявляется очень ярко. Среднее значение коэффициента корреляции по многолетним данным составляет 0,65, причем эта величина довольно устойчива в отдельные годы исследований.
Хорошо выражена и высокодостоверна, как уже указывалось, коррелятивная связь между максимальной яйценоскостью маток непосредственно перед началом главного медосбора (в условиях средней полосы обычно в конце июня) и медопродуктивностью.
Коэффициент корреляции между этими признаками в среднем за 5 лет составил 0,50 с незначительными колебаниями по годам.
Яйценоскость пчелиных маток в решающей мере определяет жизнедеятельность пчелиной семьи в целом. Этот важнейший хозяйственно полезный признак может рассматриваться, как говорилось выше, и как самостоятельный селективный, имеющий определяющее значение при разведенческой специализации пчеловодства, и как косвенный — при селекции на другие признаки, в частности на медопродуктивность.
У пчел среднерусской породы в условиях среднелетнего главного медосбора связь между среднесуточной яйценоскостью за 36 дней до начала главного медосбора и медопродуктивностью проявляется очень ярко. Среднее значение коэффициента корреляции по многолетним данным составляет 0,65, причем эта величина довольно устойчива в отдельные годы исследований.
Хорошо выражена и высокодостоверна, как уже указывалось, коррелятивная связь между максимальной яйценоскостью маток непосредственно перед началом главного медосбора (в условиях средней полосы обычно в конце июня) и медопродуктивностью.
Коэффициент корреляции между этими признаками в среднем за 5 лет составил 0,50 с незначительными колебаниями по годам.
Яйценоскость, подсчитанная в конце июня, объективно характеризует потенциальные возможности маток. Между среднесуточной яйценоскостью в конце июня и среднесуточной яйценоскостью за 36 дней до главного медосбора существует высокая положительная коррелятивная связь. Среднее значение коэффициента корреляции по многолетним данным составляет 0,82. Таким образом, яйценоскость маток, подсчитанную в конце июня, можно использовать для экспресс-оценки этого признака вообще.Среднерусские пчелиные матки вслед за итальянскими являются высокояйценоскими. Это дает возможность пчелиным семьям среднерусской породы нарастить большую массу пчел для использования медосбора, а также к зимовке. Однако для полного проявления этого признака, кроме создания оптимальных условий ухода и содержания необходимо проведение целенаправленной селекционной работы. Без соблюдения этих условий потенциальные возможности среднерусских пчелиных маток могут остаться нереализованными.
В селекционной работе 1970—1983 гг. отбор на медопродуктивность пчелиных семей среднерусской породы сочетали с отбором на яйценоскость маток. При этом генотипический отбор проводили только по женской линии, по мужской линии отбор был массовым.
Характеристика отбираемых на племя (для репродукции очередной генерации) пчелиных семей по среднесуточной яйценоскости их маток приведена в табл. 14.
 Абсолютные показатели яйценоскости племенных маток сравниваются с показателями маток-сверстниц из рядовых пчелиных семей, не подвергавшихся систематическому отбору, а также с показателями маток последовательных генераций выводимой линии.
Абсолютные показатели яйценоскости племенных маток сравниваются с показателями маток-сверстниц из рядовых пчелиных семей, не подвергавшихся систематическому отбору, а также с показателями маток последовательных генераций выводимой линии.
Из приведенных в табл. 14 данных о среднесуточной яйценоскости этих же племенных пчелиных семей видно, что показатели яйценоскости маток исходной популяции были невысокими. Для репродукции первого поколения была отобрана матка, имевшая среднесуточную яйценоскость в среднем за 36 дней до начала главного медосбора всего лишь 1256 яиц.
Этот первый отбор дал возможность для репродукции F-2 найти матку с рекордной среднесуточной яйце носкостью — 2256 яиц в сутки. Для репродукции последующих поколений были использованы матки с достаточно высокой яйценоскостью, хотя она была ниже яйценоскости родоначальницы F-2.
За все годы отбора среднесуточная яйценоскость за 36 дней до начала главного медосбора у отбираемых маток была значительно выше (в среднем на 49,2%) яйценоскости маток-сверстниц, а превосходство над среднелинейными показателями было значительно меньшим (14,2%) в результате усиливающейся консолидации селекционируемой линии по этому признаку.
Проводимый отбор вызвал существенное повышение уровня яйценоскости маток (см. табл. 15). Уровень среднесуточной яйценоскости маток исходной популяции в последующих поколениях был значительно превышен.

Из приведенных в табл. 14 данных о среднесуточной яйценоскости этих же племенных пчелиных семей видно, что показатели яйценоскости маток исходной популяции были невысокими. Для репродукции первого поколения была отобрана матка, имевшая среднесуточную яйценоскость в среднем за 36 дней до начала главного медосбора всего лишь 1256 яиц.
Этот первый отбор дал возможность для репродукции F-2 найти матку с рекордной среднесуточной яйце носкостью — 2256 яиц в сутки. Для репродукции последующих поколений были использованы матки с достаточно высокой яйценоскостью, хотя она была ниже яйценоскости родоначальницы F-2.
За все годы отбора среднесуточная яйценоскость за 36 дней до начала главного медосбора у отбираемых маток была значительно выше (в среднем на 49,2%) яйценоскости маток-сверстниц, а превосходство над среднелинейными показателями было значительно меньшим (14,2%) в результате усиливающейся консолидации селекционируемой линии по этому признаку.
Проводимый отбор вызвал существенное повышение уровня яйценоскости маток (см. табл. 15). Уровень среднесуточной яйценоскости маток исходной популяции в последующих поколениях был значительно превышен.

Для выявления селекционного эффекта каждое поколение сравнивалось с контрольным, в котором были пчелиные семьи с матками-сверстницами, не подвергавшимися целенаправленному отбору.
Наибольший селекционный эффект (1977 яиц в сутки, почти в 2 раза больше исходного уровня) получен в F-4.
Фенотипическая изменчивость яйценоскости маток выводимой линии была несколько ниже, чем в контроле, а в F6 по сравнению с исходной популяцией Cv% снизился на 10,3%. Такое снижение фенотипической изменчивости можно считать следствием отбора, хотя более объективен, на наш взгляд, был бы анализ генотипической изменчивости.
Характеристика максимальной яйценоскости племенных среднерусских маток, развиваемой ими обычно в конце июня перед началом главного медосбора, приведена в табл. 16. Изменение ее в процессе отбора (табл. 17) показывает, что наибольший уровень максимальной яйценоскости достигнут также в F-4. Можно отметить также ненаправленное колебание коэффициентов фенотипической вариации.
Наибольший селекционный эффект (1977 яиц в сутки, почти в 2 раза больше исходного уровня) получен в F-4.
Фенотипическая изменчивость яйценоскости маток выводимой линии была несколько ниже, чем в контроле, а в F6 по сравнению с исходной популяцией Cv% снизился на 10,3%. Такое снижение фенотипической изменчивости можно считать следствием отбора, хотя более объективен, на наш взгляд, был бы анализ генотипической изменчивости.
Характеристика максимальной яйценоскости племенных среднерусских маток, развиваемой ими обычно в конце июня перед началом главного медосбора, приведена в табл. 16. Изменение ее в процессе отбора (табл. 17) показывает, что наибольший уровень максимальной яйценоскости достигнут также в F-4. Можно отметить также ненаправленное колебание коэффициентов фенотипической вариации.

Таким образом, яйценоскость пчелиных маток, является важным селективным признаком, имеющим как самостоятельную селекционную значимость, так и пригодным для проведения косвенного отбора на медопродуктивность. Уже при жестком отборе можно довести уровень яйценоскости до предельных значений, превышающих стандарт породы.
 В табл. 18 обобщен селекционный эффект, полученный по всем селективным признакам. Данные показывают, что в среднем за 9 поколений селекции превышение над уровнем исходной популяции составило по медопродуктивности 57,9%, по максимальной яйценоскости 40,2% и по среднесуточной — 64,4%. Превосходство над уровнем селективных признаков по сравнению с матками-сверстницами несколько ниже (28,3—41,9%), причем к F-7 оно становится незначительным (4,2—5,0%). Очевидно, до ничтожно малых значений снизился уровень генотипического разнообразия признаков. Это же подтверждает и анализ степени наследуемости селективных признаков. Их значения с 8— 26% в F-1 достигли 1—3% в F-7. Значительно снизились и значения коэффициентов фенотипической и генотипической изменчивости.
В табл. 18 обобщен селекционный эффект, полученный по всем селективным признакам. Данные показывают, что в среднем за 9 поколений селекции превышение над уровнем исходной популяции составило по медопродуктивности 57,9%, по максимальной яйценоскости 40,2% и по среднесуточной — 64,4%. Превосходство над уровнем селективных признаков по сравнению с матками-сверстницами несколько ниже (28,3—41,9%), причем к F-7 оно становится незначительным (4,2—5,0%). Очевидно, до ничтожно малых значений снизился уровень генотипического разнообразия признаков. Это же подтверждает и анализ степени наследуемости селективных признаков. Их значения с 8— 26% в F-1 достигли 1—3% в F-7. Значительно снизились и значения коэффициентов фенотипической и генотипической изменчивости.Из этих данных можно сделать вывод о том, что такие признаки, как медопродуктивность и особенно яйценоскость среднерусских маток, отзывчивы на селекцию. Наибольший эффект отмечен в F-1—F-4. Очевидно,в селекции среднерусских пчел целесообразно иметь дело с короткими линиями (до F-1—F-4).
 Одним из аспектов селекционной работы может быть селекция линий среднерусских пчел, специализированных на опылении отдельных энтомофильных культур, в частности гречихи.
Одним из аспектов селекционной работы может быть селекция линий среднерусских пчел, специализированных на опылении отдельных энтомофильных культур, в частности гречихи.Флоромиграцию пчелиных определяют цвет, запах цветков, наличие нектара и пыльцы и их доступность. Поскольку нектар растений различных семейств отличается по составу сахаров, а также по аминокислотному составу, то эти факторы также могут влиять на флоромиграцию и играть определенную роль в коэволюции растений и опылителей (Харборн, 1985).
У одиночных пчел наблюдается довольно узкая приспособленность к растениям определенных семейств. Так, представители рода Andrene посещают в основном орхидные.
Общественные пчелы, и в частности Apis mellifera, посещают довольно широкий спектр энтомофильных растений, отдавая предпочтение семействам губоцветных, норичниковых и бобовых, цветки которых имеют главным образом голубой и желтый цвета. Однако и внутри вида медоносной пчелы отмечается избирательность различных пород (рас) пчел по отношению к тем или иным растениям. Еще И.Н.Клинген (1912) убедительно показал преимущество длиннохоботных серых горных кавказских пчел в опылении красного клевера.
Ш.О.Гасанов (1970), подтверждая данные предыдущих исследований о наличии флороспециализации у пчел разных пород (рас), показал, что среднерусские пчелы предпочитают гречиху и липу, серые горные кавказские— клевера, сурепку, фацелию.
Принимая наличие флороспециализации как бесспорный факт, селекционеры стали предпринимать попытки выведения линий пчел, специализированных на опылении отдельных культур: люцерны (Макензен и Най, 1967; Cale, 1971; И.К.Давыденко и др., 1980), красного и белого клеверов (Холм, 1971).
Совместно с Д.В.Гирником мы изучили возможность селекции пчел на опыление гречихи. Наши исследования косвенно подтверждают вывод: пчелы, отловленные на цветках гречихи, по своим экстерьерным признакам ближе к стандарту среднерусской породы, чем пчелы, посещающие цветки других растений (табл. 19).

Возникает необходимость усиления опылительной деятельности пчел на этой культуре.
Уже подчеркивалось, что выбор главного селективного признака имеет определяющее значение. Известно, что с одних растений пчелы той или иной породы предпочитают сбор нектара, с других — сбор пыльцы, с третьих — одинаково хорошо собирают и пыльцу и нектар одновременно. Если пчелы испытываемой породы охотно собирают пыльцу с данной культуры, то в качестве основного селективного признака следует брать именно его. Если же они охотно собирают и пыльцу и нектар, то тоже лучше брать в качестве основного селективного признака склонность собирать пыльцу, так как провести ботанический анализ пыльцы проще, чем сделать подобный анализ нектара.
Мы проследили за работой на цветках гречихи пчел, собирающих пыльцу, и пчел, собирающих нектар, а также учли завязываемость семян гречихи после опыления их пыльце- и нектаро сборщицами. Эффективность опыления цветков гречихи пчелами разной специализации составила (%):
Пыльценектаросборщицы (54 пчелы) ........... 78,3 (94 завязавшихся зерна)
Нектаросборщицы (29 пчел) ............... .21,7 (26 завязавшихся зерен)
Всего (83 пчелы) .................... 100 (120 завязавшихся зерен)
Около 80% опылительной работы на цветках гречихи приходится на пчел, собирающих одновременно и пыльцу и нектар, т. е. на пыльце-нектаро сборщиц. Полученные данные указывают на то, что в качестве главного селективного признака при выведении специализированных («гречишных») линий, следует использовать склонность отдельных семей пчел среднерусской породы собирать пыльцу этой культуры.
Чтобы изучить склонность той или иной пчелиной семьи собирать пыльцу с гречихи, на каждую семью испытываемой популяции за неделю до начала цветения гречихи навешивали пыльцеуловитель. Заблаговременное навешивание пыльцеуловителей позволило пчелам привыкнуть летать через пыльцеуловитель с отключенной пыльцеотбирающей решеткой.
Обножки у пчел отбирали трехкратно в течение периода массового цветения. При большом количестве испытываемых семей (300—500 и более) пыльцеуловители можно включать однократно, так как данные однократного отбора довольно высоко коррелируют как со смежными отборами (г=0,5 — 0,7), так и со средней величиной этого показателя за сезон (г=0,6 — 0,8). Поскольку характер корреляций зависит от многих причин, их следует рассчитывать в каждом конкретном случае.
Лучшим временем суток для включения пыльцеуловителей является период с 8 до 12 ч, т.е. время наилучшего лета пчел на гречиху. Если погода недостаточно благоприятна для этого, то включение и выключение пыльцеуловителей переносится на более позднее время (с 9 до 13 или даже с 10 до 14 ч).
После того как пыльцеуловители отключены (на включение или выключение 100 пыльцеуловителей один человек затрачивает 3—5 мин), обножки из приемных лотков пыльцеуловителей осторожно пересыпают в сухие бумажные пакеты, на которых отмечают дату сбора и номер семьи.
До начала ботанического анализа пакеты с пыльцой хранят в сухом, защищенном от прямых солнечных лучей месте. Как только обножки немного подсохнут (станут сыпучими), проводят их ботанический анализ. Для этого обножки каждой пробы на чистом белом листе бумаги с помощью шпателя разбирают на фракции по их цвету. Если простым глазом затруднительно отделить обножки пыльцы одних растений от других, то разбор производят с помощью лупы.
Ботаническую принадлежность каждой фракции обножек определяют под микроскопом. Контроль за правильностью определения ботанической принадлежности обножек осуществляют с помощью постоянных препаратов пыльцы.
В процессе работы важно определить значения генетико-популяционных параметров селективного признака.
В наших исследованиях в разные годы пределы варьирования «гречишности» составляли 12—68%, q была на уровне 15,8—21,0%, а коэффициент вариации колебался в пределах 42,5—86,0%. Такие показатели фенотипической изменчивости «гречишности» дают основание надеяться на успех селекции специализированных «гречишных» линий среднерусских пчел.
Как известно, повторяемость признака является в определенном смысле верхним пределом его наследуемости. Для получения этого показателя рассчитывают коэффициенты корреляции между смежными наблюдениями данного сезона, а также их квадраты. Результаты наших исследований за 4 года показали, что доля действия общих факторов, определяющих проявление селективного признака «гречишности», довольно высока и составляет в разные годы 24—25%. Такая сравнительно высокая повторяемость признака «гречишности» дает основание рассчитывать на его селективное постоянство.
Ниже приведена повторяемость (%) приноса гречишной пыльцы, собранной пчелами в течение сезона (П = 33 — 67):
Год наблюдений
(количество сопоставимых учетов) r±mT r2
1. й (5) 0,47 ±0,040 0,24
2. й (3) 0,67 ±0,068 0,45
3. й (3) 0,61 ±0,064 0,37
4. й (3) 0,49 ±0,131 0,27
На уровень постоянства признака «гречишности» указывает также характер корреляции между процентом гречишной пыльцы, приносимой пчелами за отдельные учеты, и средней величиной этого признака за все учеты сезона. Наши четырехлетние данные показывают высокую и устойчивую положительную корреляцию между этими признаками (г=0,68 — 0,87).
Таким образом, даже разовые учеты процента гречишной пыльцы в общем сборе могут дать достаточно объективное представление о склонности пчелиной семьи к опылительной работе на гречихе.
Склонность пчелиных семей к сбору пыльцы с гречихи можно выявить также путем расчета коэффициента корреляции при сопоставлении данных по одним и тем же пчелиным семьям за разные годы. В наших исследованиях значения коэффициентов корреляции колебались от 0,43±0,144 до 0,73±0,81. В целом данные об уровне повторяемости признака склонности пчелиных семей собирать пыльцу с гречихи как в течение сезона, так и в разные годы, свидетельствуют о его надежности в селекционном смысле и дают возможность проводить его оценку даже в течение одного сезона и по отдельным учетам. Это позволяет значительно сократить сроки селекционного процесса выведения специализированных линий среднерусских пчел для опыления гречихи.
Известно, что наследуемость есть доля наследственной обусловленности изменчивости признака в популяции. Степень наследуемости признака характеризуется коэффициентом h2, величина которого зависит от генетической структуры популяции, изменчивости признака и условий внешней среды. Показатель наследуемости служит одним из критериев эффективности отбора и позволяет прогнозировать результаты селекции.
В нашей работе при строго контролируемом инструментальном осеменении коэффициент наследуемости признака «гречишности» равнялся 0,25. Это говорит о том, что изменчивость признака «гречишности» в испытываемой популяции среднерусских пчел на 25% обусловлена генетически.
По полученным данным можно сделать прогноз эффекта селекции на одно поколение. Для этого используют формулу: R — Sh2, где R — ответ на селекцию; 5 — селекционный дифференциал, определяющий жесткость отбора; h2 — коэффициент наследуемости.
Используя данные наших исследований, мы составили следующий прогноз на одно поколение.
Оказалось, что если в племенную группу отобрать пчелиные семьи, собирающие 49,4% гречишной пыльцы при среднепопуляционном значении этого показателя 35,2% и коэффициенте наследуемости, равном 0,25, то семьи с выведенными матками от племенных семей будут собирать 38,7% гречишной пыльцы (35,2+3,5) при S = 14,2 (49,4—35,2), так как ответ на селекцию (R) будет равен 3,5% (14,2*0,25).
Характер наследования склонности среднерусских пчел собирать пыльцу с гречихи прослеживается на примере результатов наших исследований. На Орловской опытной станции нами была испытана популяция среднерусских пчел на склонность семей собирать пыльцу с гречихи. По результатам этих испытаний были выделены 4 семьи. Одна пара родительских семей имела относительно высокую склонность собирать пыльцу с гречихи (54,8 и 54,9%), другая родительская пара — относительно низкую (13,9 и 30,0%).
Среднепопуляционный уровень признака составлял 38,1% при Cv=42,5%. Осеменение маток спермой трутней от подобранных семей производили инструментально. Были испытаны по селективному признаку и полученные группы дочерей.
Приведенные ниже данные содержания (%) гречишной пыльцы в общем сборе позволяют выявить характер наследования склонности среднерусских пчел к этой культуре:
Обножки у пчел отбирали трехкратно в течение периода массового цветения. При большом количестве испытываемых семей (300—500 и более) пыльцеуловители можно включать однократно, так как данные однократного отбора довольно высоко коррелируют как со смежными отборами (г=0,5 — 0,7), так и со средней величиной этого показателя за сезон (г=0,6 — 0,8). Поскольку характер корреляций зависит от многих причин, их следует рассчитывать в каждом конкретном случае.
Лучшим временем суток для включения пыльцеуловителей является период с 8 до 12 ч, т.е. время наилучшего лета пчел на гречиху. Если погода недостаточно благоприятна для этого, то включение и выключение пыльцеуловителей переносится на более позднее время (с 9 до 13 или даже с 10 до 14 ч).
После того как пыльцеуловители отключены (на включение или выключение 100 пыльцеуловителей один человек затрачивает 3—5 мин), обножки из приемных лотков пыльцеуловителей осторожно пересыпают в сухие бумажные пакеты, на которых отмечают дату сбора и номер семьи.
До начала ботанического анализа пакеты с пыльцой хранят в сухом, защищенном от прямых солнечных лучей месте. Как только обножки немного подсохнут (станут сыпучими), проводят их ботанический анализ. Для этого обножки каждой пробы на чистом белом листе бумаги с помощью шпателя разбирают на фракции по их цвету. Если простым глазом затруднительно отделить обножки пыльцы одних растений от других, то разбор производят с помощью лупы.
Ботаническую принадлежность каждой фракции обножек определяют под микроскопом. Контроль за правильностью определения ботанической принадлежности обножек осуществляют с помощью постоянных препаратов пыльцы.
В процессе работы важно определить значения генетико-популяционных параметров селективного признака.
В наших исследованиях в разные годы пределы варьирования «гречишности» составляли 12—68%, q была на уровне 15,8—21,0%, а коэффициент вариации колебался в пределах 42,5—86,0%. Такие показатели фенотипической изменчивости «гречишности» дают основание надеяться на успех селекции специализированных «гречишных» линий среднерусских пчел.
Как известно, повторяемость признака является в определенном смысле верхним пределом его наследуемости. Для получения этого показателя рассчитывают коэффициенты корреляции между смежными наблюдениями данного сезона, а также их квадраты. Результаты наших исследований за 4 года показали, что доля действия общих факторов, определяющих проявление селективного признака «гречишности», довольно высока и составляет в разные годы 24—25%. Такая сравнительно высокая повторяемость признака «гречишности» дает основание рассчитывать на его селективное постоянство.
Ниже приведена повторяемость (%) приноса гречишной пыльцы, собранной пчелами в течение сезона (П = 33 — 67):
Год наблюдений
(количество сопоставимых учетов) r±mT r2
1. й (5) 0,47 ±0,040 0,24
2. й (3) 0,67 ±0,068 0,45
3. й (3) 0,61 ±0,064 0,37
4. й (3) 0,49 ±0,131 0,27
На уровень постоянства признака «гречишности» указывает также характер корреляции между процентом гречишной пыльцы, приносимой пчелами за отдельные учеты, и средней величиной этого признака за все учеты сезона. Наши четырехлетние данные показывают высокую и устойчивую положительную корреляцию между этими признаками (г=0,68 — 0,87).
Таким образом, даже разовые учеты процента гречишной пыльцы в общем сборе могут дать достаточно объективное представление о склонности пчелиной семьи к опылительной работе на гречихе.
Склонность пчелиных семей к сбору пыльцы с гречихи можно выявить также путем расчета коэффициента корреляции при сопоставлении данных по одним и тем же пчелиным семьям за разные годы. В наших исследованиях значения коэффициентов корреляции колебались от 0,43±0,144 до 0,73±0,81. В целом данные об уровне повторяемости признака склонности пчелиных семей собирать пыльцу с гречихи как в течение сезона, так и в разные годы, свидетельствуют о его надежности в селекционном смысле и дают возможность проводить его оценку даже в течение одного сезона и по отдельным учетам. Это позволяет значительно сократить сроки селекционного процесса выведения специализированных линий среднерусских пчел для опыления гречихи.
Известно, что наследуемость есть доля наследственной обусловленности изменчивости признака в популяции. Степень наследуемости признака характеризуется коэффициентом h2, величина которого зависит от генетической структуры популяции, изменчивости признака и условий внешней среды. Показатель наследуемости служит одним из критериев эффективности отбора и позволяет прогнозировать результаты селекции.
В нашей работе при строго контролируемом инструментальном осеменении коэффициент наследуемости признака «гречишности» равнялся 0,25. Это говорит о том, что изменчивость признака «гречишности» в испытываемой популяции среднерусских пчел на 25% обусловлена генетически.
По полученным данным можно сделать прогноз эффекта селекции на одно поколение. Для этого используют формулу: R — Sh2, где R — ответ на селекцию; 5 — селекционный дифференциал, определяющий жесткость отбора; h2 — коэффициент наследуемости.
Используя данные наших исследований, мы составили следующий прогноз на одно поколение.
Оказалось, что если в племенную группу отобрать пчелиные семьи, собирающие 49,4% гречишной пыльцы при среднепопуляционном значении этого показателя 35,2% и коэффициенте наследуемости, равном 0,25, то семьи с выведенными матками от племенных семей будут собирать 38,7% гречишной пыльцы (35,2+3,5) при S = 14,2 (49,4—35,2), так как ответ на селекцию (R) будет равен 3,5% (14,2*0,25).
Характер наследования склонности среднерусских пчел собирать пыльцу с гречихи прослеживается на примере результатов наших исследований. На Орловской опытной станции нами была испытана популяция среднерусских пчел на склонность семей собирать пыльцу с гречихи. По результатам этих испытаний были выделены 4 семьи. Одна пара родительских семей имела относительно высокую склонность собирать пыльцу с гречихи (54,8 и 54,9%), другая родительская пара — относительно низкую (13,9 и 30,0%).
Среднепопуляционный уровень признака составлял 38,1% при Cv=42,5%. Осеменение маток спермой трутней от подобранных семей производили инструментально. Были испытаны по селективному признаку и полученные группы дочерей.
Приведенные ниже данные содержания (%) гречишной пыльцы в общем сборе позволяют выявить характер наследования склонности среднерусских пчел к этой культуре:

Мы провели индивидуальный отбор пчелиных семей из группы с высокой склонностью к сбору гречишной пыльцы и из группы с низкой склонностью и вывели от них второе поколение маток. Индивидуальный отбор пчелиных семей для вывода трутней по данному селективному признаку не проводили, и спаривание было свободным. И в этом случае имело место наследование склонности к сбору гречишной пыльцы. Однако разница между указанными группами была значительно меньше и составляла лишь 7%. Полученные данные говорят о том, что при имеющемся уровне наследуемости (/г2=0,25) массовый фенотипический отбор на «гречишность» имеет определенный смысл, но для получения большего эффекта необходимо применять генотипический отбор и подбор, что возможно только при условии строгого контроля над спариванием..
Таким образом, было подтверждено наличие у среднерусских пчел флороспециализации к гречихе и показана принципиальная возможность селекции линий, специализированных на опылении этой культуры.
Принципиальная схема чистопородной селекции
На основании изученной литературы и проведенных исследований можно предложить следующую примерную схему чистопородной селекции с породами пчел.
Ознакомление с генофондом и ресурсами породы, выявление в результате обработки литературы, анкетирования, а также экспедиционных поисков сохранившихся в чистоте в пределах ареала естественного обитания чистопородных пчел (популяции, экотипы, отдельные пасеки). Уточнение по комплексу экстерьерных, хозяйственно полезных, биологических признаков их породной принадлежности. Изучение имеющейся у пчеловода документации относительно происхождения и продуктивности пчелиных семей. Завоз маток (желательно одновозрастных) или пчелопакетов, пчелиных семей из выявленных пунктов в научно-исследовательское или опытное учреждение по пчеловодству. Организация на месте достаточно больших групп пчелиных семей нужного происхождения с использованием инструментального осеменения или изолированных случных пунктов (3—4 года).
Сравнительное испытание завезенных из разных мест популяционных групп пчелиных семей по комплексу хозяйственно полезных и биологических признаков. Выделение для дальнейшей работы перспективных популяций, обладающих комплексом ценных хозяйственно полезных признаков и пригодных для закладки на их основе заводских линий (представляют интерес и популяции, характеризующиеся каким-либо одним ценным качеством для закладки специализированной линии). Установление делового контакта с учреждениями и пчеловодами-любителями тех мест, из которых были завезены пчелы, оказавшиеся лучшими в сравнительном испытании, с целью осуществления повторных завозов для «освежения крови». Проведение необходимых мероприятий, препятствующих метизации аборигенных пчел в этих местах (создание заказников, заповедных зон и т. д ) (2—3 года).
Размножение пчелиных семей лучших популяций (желательно в условиях пространственной изоляции), организация популяционных пасек. Осуществление контроля над спариванием известными методами (изоляция в пространстве и времени, применение инструментального осеменения, двукратная смена маток). Репродуцирование наиболее чистопородного материала в центре популяционного массива, где также осуществляется репродукция маток как для расширения массива, так и для реализации на сторону.
Проведение в популяционных массивах постоянной работы по селекционному улучшению и контролю за чистопородностью, фенотипическая оценка пчелиных семей популяционных пасек, отбор предполагаемых родоначальниц линий и получение от них групп маток-дочерей для проверки их по качеству потомства. Анализ фенотипической и генотипической изменчивости, наследуемости, повторяемости, предварительный прогноз эффекта селекции. Изучение комплекса корреляционных связей между хозяйственно полезными и биологическими признаками с целью применения косвенного отбора, играющего особо важную роль в селекционном улучшении признаков с низкой наследуемостью.
Выведение аутбредных (по типу заводских) линий пчел, характеризующихся четко выраженным развитием основных селективных признаков (медопродуктивности, яйценоскости) при сохранении других хозяйственно полезных признаков на уровне стандарта для породы. Продолжение линий в 3—4 поколениях, анализ генетико-популяционных параметров, выяснение целесообразности получения последующих поколений и закладки новых линий.
Получение товарных внутрипородных гибридов (межпопуляционных и межлинейных) с целью хозяйственного использования эффекта микрогетерозиса. Испытание полученных гибридов на сочетаемость с целью последующего использования лучших вариантов в промышленных внутрипородных скрещиваниях.
Создание сети пчелоразведенческих хозяйств для массовой репродукции маток, установление обратной связи с заказчиками с целью корректировки селекционированного процесса и вовлечения в него ценного материала.
Важно подчеркнуть, что параллельно с селекционной работой мы осуществляли первичную репродукцию племенных маток линии № 44 для зоны районирования среднерусской породы.

На Орловской опытной станции массив этой линии составляет 780 пчелиных семей.
За годы работы (1976—1989) в областях районирования (Орловской, Курской, Липецкой, Брянской, Тульской, Московской, Ленинградской, Рязанской, Воронежской, Кировской, Калининградской, Псковской, Пермской, Курганской, Иркутской, Омской, Челябинской, Оренбургской, Свердловской, Костромской, а также в Алтайском крае) было реализовано около 80 тыс. неплодных и около 10 тыс. плодных маток линии № 44.
Среднегодовая прибавка медопродуктивности за период селекции 1973—1988 гг. за счет селекционного улучшения в расчете на одну семью пчел составила 11,2 кг, или, в денежном выражении, за вычетом дополнительных затрат на селекционный процесс — 35,99 руб (в ценах 1990 г.).
Важно подчеркнуть, что параллельно с селекционной работой мы осуществляли первичную репродукцию племенных маток линии № 44 для зоны районирования среднерусской породы.

На Орловской опытной станции массив этой линии составляет 780 пчелиных семей.
За годы работы (1976—1989) в областях районирования (Орловской, Курской, Липецкой, Брянской, Тульской, Московской, Ленинградской, Рязанской, Воронежской, Кировской, Калининградской, Псковской, Пермской, Курганской, Иркутской, Омской, Челябинской, Оренбургской, Свердловской, Костромской, а также в Алтайском крае) было реализовано около 80 тыс. неплодных и около 10 тыс. плодных маток линии № 44.
Среднегодовая прибавка медопродуктивности за период селекции 1973—1988 гг. за счет селекционного улучшения в расчете на одну семью пчел составила 11,2 кг, или, в денежном выражении, за вычетом дополнительных затрат на селекционный процесс — 35,99 руб (в ценах 1990 г.).
Фактический объем внедрения плодных маток линии № 44 составил 10 тыс. шт., и следовательно, экономический эффект, выраженный через прибавку медопродуктивности, равен 3599 тыс. руб. По нашим расчетам, с расширением объема репродукции до удовлетворения полного спроса эффект может составить около 26 млн руб. (в ценах 1990 г.).
ИСПОЛЬЗОВАНИЕ СРЕДНЕРУССКИХ ПЧЕЛ В МЕЖПОРОДНЫХ ПОМЕСЯХ
В настоящее время можно считать бесспорно доказанным факт проявления гетерозиса при скрещивании пчел некоторых завозных пород с местной среднерусской (А.Н.Брюханенко, 1929; А.А.Курочкин, 1030; М.3.Краснопеев по В.В.Алпатову, 1948 Г.Д.Билаш, 1956; Г.Ф.Таранов, 1956; А.Н.Биктимиров, 1963; А.Я.Шекшуев, 1967; А.В.Бородачев, В.Т.Бородачева, 1982, и др.).
Перечисленные авторы, как правило, использовали маток завозной породы, а трутней — местной среднерусской. Полученные помеси наследовали такие качества исходных пород, как хорошая зимостойкость, кормодобывающая активность, работоспособность. Эффект гетерозиса проявлялся на качестве рабочих пчел. Вместе с тем при простых скрещиваниях генотип матки не затрагивается, и, следовательно, эффект гетерозиса по яйценоскости маток, а также улучшающее действие другой породы не проявляются.
В то же время матки среднерусской породы обладают высоким потенциалом яйценоскости, который можно было бы использовать в результате скрещивания для усиления этого признака у пород пчел с низкой яйценоскостью.
Помесными матками будут дочери маток от простого скрещивания, спарившиеся с трутнями другой породы, если же их спарить с трутнями третьей породы, рабочие пчелы от них будут сложными трехкратными помесями. Впрочем, помесными будут любые матки-дочери простых помесей. Но если не ставится цель использования в генотипе рабочих пчел третьей породы, можно провести возвратное скрещивание с трутнями одной из уже использованных пород, что повлечет за собой увеличение доли «кровности» этой породы в генотипе пчел до 75%.
Подобные сложные (трехкратные) межпородные помеси с участием среднерусской породы были испытаны в 1979—1983 гг. в Краснодарском крае (Майкопский и Краснополянский опорные пункты пчеловодства) и Башкирии (Башкирская опытная станция пчеловодства). Кроме среднерусской породы (Ср) в помесях использовали пчел кавказской (Кав), итальянской (Ит), карпатской (Кар), краинской (Кр) пород и дальневосточных пчел (Дв). Пчелы среднерусской породы являлись обязательным компонентом сложных помесей. В качестве контрольных были использованы местные породы пчел (серая горная кавказская на юге и среднерусская в Башкирии).
Контроль над спариванием при получении сложных помесей осуществляли или методом инструментального осеменения в НИИ пчеловодства, или в результате использования мощного трутневого барьера в зонах чистопородного разведения на местах.
Среди прочих задач было и выяснение влияния высокояйценоских пород пчел (в частности, среднерусской) на генотип поместных маток с целью увеличения их яйценоскости и, соответственно, медопродуктивности.
В табл. 20 приведены данные о среднесуточной яйценоскости маток перед началом главного медосбора.

пчеловодства (ОПП) отмечено очевидное превосходство помесных маток над серыми горными кавказскими, наиболее высокая яйценоскость зафиксирована в варианте ИтХ ХСрХКав за счет высокой яйценоскости двух пород (итальянской и среднерусской), определяющих генотип маток.
На Башкирской ОСП в лесостепной зоне наблюдалось некоторое превосходство (12,2% в среднем за два года) помесных маток варианта ИтХКарХСр над среднерусскими за счет положительного влияния высокояйценоской итальянской породы, а в условиях степи в 1983 г. помесные матки без участия в их генотипе среднерусской породы даже не смогли сравняться в яйценоскости с чистопородными среднерусскими.
На Майкопском ОПП было выявлено убедительное превосходство яйценоскости помесных маток с участием среднерусской и итальянской пород над чистопородными серыми горными кавказскими, хотя доля «кровности» среднерусской породы в генотипе маток в 1980 г. составляла всего лишь 25%.
В табл. 21 показана сила пчелиных семей с матками-помесями на медосборе. Здесь также хорошо прослеживается положительное влияние наследственности среднерусской и итальянской высокояйценоских пород.

На Краснополянском ОПП сила семей с помесными матками, имеющими в 1979 г. 50% наследственности среднерусской породы, была на 20—56% выше, чем в контрольной группе. В 1980 г. в результате возвратного скрещивания с трутнями серой горной кавказской породы доля «кровности» среднерусской породы снизилась до 25%, что повлекло и снижение силы пчелиных семей.
На Башкирской ОСП на силу пчелиных семей влияло участие в генотипе помесных маток итальянской породы. Это влияние было положительным, но незначительным, так как среднерусские пчелиные семьи в контрольной группе сами в силу своих породных особенностей склонны к наращиванию значительной силы к медосбору.
На Майкопском ОПП влияние среднерусской породы на силу пчелиных семей было положительным и очевидным даже при доле «кровности» этой породы 25%. Положительно влияла и итальянская порода, доля «кровности» которой составила 50%, что обеспечило увеличение силы пчелиных семей на 43,2—58,3% по сравнению с контрольными.
На медопродуктивность сложных межпородных помесей (табл. 22) оказывали влияние как яйценоскость маток и сила пчелиных семей, так, очевидно, и повышенная работоспособность рабочих пчел за счет гетерозиса.
На Башкирской ОСП на силу пчелиных семей влияло участие в генотипе помесных маток итальянской породы. Это влияние было положительным, но незначительным, так как среднерусские пчелиные семьи в контрольной группе сами в силу своих породных особенностей склонны к наращиванию значительной силы к медосбору.
На Майкопском ОПП влияние среднерусской породы на силу пчелиных семей было положительным и очевидным даже при доле «кровности» этой породы 25%. Положительно влияла и итальянская порода, доля «кровности» которой составила 50%, что обеспечило увеличение силы пчелиных семей на 43,2—58,3% по сравнению с контрольными.
На медопродуктивность сложных межпородных помесей (табл. 22) оказывали влияние как яйценоскость маток и сила пчелиных семей, так, очевидно, и повышенная работоспособность рабочих пчел за счет гетерозиса.

На Краснополянском ОПП на фоне относительно скудного медосбора все же наблюдалось превосходство по этому признаку помесей с участием среднерусской породы над чистопородными пчелиными семьями серой горной кавказской породы.
На Башкирской ОСП превосходство по медопродуктивности сложных помесей над контрольной группой в обеих зонах было убедительным и значительным.
На Башкирской ОСП превосходство по медопродуктивности сложных помесей над контрольной группой в обеих зонах было убедительным и значительным.
Что же касается Майкопского ОПП, то наиболее продуктивными оказались помеси с долей «кровности» среднерусской породы в генотипе рабочих пчел 50%; в 1982 г. эти пчелиные семьи собрали в среднем по 64,2 кг меда, что для местных условий можно считать значительным достижением.
Изучение характера корреляций между такими признаками помесей, как сила пчелиных семей на начало главного медосбора и их медопродуктивность, представляет интерес. Мы изучали этот вопрос в процессе испытания различных вариантов межпородного скрещивания с участием среднерусских пчел, проводимого в различных природно-климатических зонах.
Полученные данные (табл. 23) показывают, что характер корреляций силы семей и медопродуктивности различен в зависимости от происхождения пчелиных семей, места и года испытаний.

В большинстве случаев выявлена положительная корреляция между этими признаками, однако в некоторых вариантах наблюдается отсутствие корреляции и даже тенденция к отрицательной зависимости. Это касается прежде всего пчел кавказской породы и помесей с их участием. Так, на Краснополянском ОПП коэффициенты корреляции у чистопородных серых горных кавказских пчел составили лишь 0,03—0,47; в испытаниях 1980 и 1982 гг., когда доля их наследственности у помесей составляла 75%, значения коэффициентов корреляции также были чрезвычайно низкими.
Это объясняется, очевидно, особенностью серых горных кавказских пчел, заключающейся в их большей предприимчивости и способности эффективно использовать даже ничтожный медосбор. В этом случае значение большой силы пчелиных семей как бы нивелируется.
Сравнительно высокие значения коэффициентов корреляции были обнаружены у среднерусских пчел и помесей с их участием на Башкирской ОСП, Приморском ОПП и Чимкентском ОПП В этом случае на характер корреляций оказывала влияние такая породная особенность среднерусских пчел, как способность эффективно использовать лишь сильный медосбор.
Сходный с этим характер корреляций отмечен у пчелиных семей дальневосточной и краинской пород в местах их районирования (Приморский край и Чимкентская область Казахстана).
Сила корреляционных связей изменяется и в зависимости от условий сезона испытания, хотя и не в такой степени, как от происхождения пчелиных семей. В типичных для данной местности условиях следует ожидать, что различия будут несущественными. См.: Семьи-помеси под контроль! Старая история о наилучшей пчеле... Экологический гетерозис пчелиных семей и ошибки породного районирования. Породное районирование и лучшие пчелы для России. О гетерозисе.
ЗАКЛЮЧЕНИЕ
Пчелы среднерусской породы благодаря выработавшимся в процессе эволюции качествам — хорошей зимостойкости, устойчивости к ряду заболеваний, способности эффективно использовать короткий, но бурный медосбор, высокой плодовитости маток — незаменимы для разведения в центральных и северных зонах страны с их неблагоприятными условиями. К сожалению, в большинстве мест своего ареала среднерусские пчелы метизированы пчелами южных пород. Восстановление генофонда породы, размножение и селекционное улучшение— важная народнохозяйственная задача.
Генофонд среднерусской породы представлен рядом популяций, отличающихся морфологическими признаками. В процессе сравнительного испытания татарской, владимирской, новосибирской, уральской, вологодской и орловской популяций наиболее перспективной оказалась татарская, превысившая местных орловских пчел по среднесуточной яйценоскости маток на 17,9%, по медопродуктивности — на 25,7% и показавшая хорошую зимостойкость.
Исследования, проведенные за несколько десятилетий НИИ пчеловодства, позволили внести уточнения в методику определения чистопородности пчел и выделения популяций по экстерьерным признакам путем анализа их изменчивости. Определен объем выборки, необходимый для получения объективных данных по экстерьерным характеристикам внутрипородных группировок.
Изучение хозяйственно полезных признаков среднерусских пчел показало значительную фенотипическую изменчивость (особенно сильно варьируют показатели зимостойкости). Значительна степень генотипической изменчивости и наследуемости признаков, что дает основание рассчитывать на достаточную эффективность отбора. Однако эти параметры не остаются постоянными, они изменяются в процессе селекции и являются индикаторами его эффективности.
Установлены достоверные положительные фенотипические корреляции с медопродуктивностью пчел некоторых признаков: размеров крыла, тергита, величины кубитального индекса, зимостойкости, среднесуточной и максимальной яйценоскости маток, общей массы пчел перед медосбором.
Показано, что зимостойкость коррелирует с яйценоскостью маток при первом весеннем учете, со среднесуточной яйценоскостью за 36 дней до начала главного медосбора и с максимальной яйценоскостью. Все признаки, определяющие зимостойкость, коррелируют между собой, а также со степенью пораженности нозематозом, активностью каталазы ректальных желез, процентом сахарной добавки в зимних кормах. С максимальной яйценоскостью маток коррелирует их среднесуточная яйценоскость, а также их масса и размеры тела. Рассчитанные корреляции открывают широкие возможности для проведения косвенного отбора по сигнальным признакам.
Среднерусские пчелы отзывчивы на отбор. В среднем за 9 поколений превосходство над исходной популяцией составило по медопродуктивности и среднесуточной яйценоскости маток 60%, по максимальной яйценоскости — 40%. Наивысшие показатели среднесуточной яйценоскости маток и их максимальной яйценоскости были достигнуты в четвертом поколении.
Показана возможность использования высокого потенциала яйценоскости для производства на пасеках 100% пчелопакетов без существенного отрицательного влияния на медопродуктивность.
Поскольку при жестком отборе для селекционной работы уже к третьему поколению снижается генотипическое разнообразие признаков, усиливается их константность, уменьшается эффективность массового отбора, то наиболее перспективны для улучшения среднерусских пчел короткие линии. Для дальнейшего развития селективных признаков целесообразно использовать методы, повышающие их генотипическое разнообразие (кроссы линий, популяций).
Пчелы среднерусской породы отличаются хорошей сочетаемостью при скрещивании с другими породами, обеспечивающей получение эффекта гетерозиса как при простых двухпородных (генотип рабочих пчел), так и при сложных трехпородных скрещиваниях, затрагивающих и генотип маток. Так, среднее превосходство 8 вариантов сложных скрещиваний с использованием пчел среднерусской породы в четырех испытательных пунктах по яйценоскости составило 20%, а по медопродуктивности — 21% по сравнению с районированной породой. Положительный эффект создается за счет зимостойкости и яйценоскости маток, влияние которых распространяется и на другие признаки.
За 1976—1990 гг. в 30 областях районирования было реализовано более 80 тыс. неплодных и около 10 тыс. плодных маток отселекционированной линии № 44. Сплошной массив этой линии на Орловской опытной станции пчеловодства составил 1017 пчелиных семей.
На основе полученных материалов была организована и оформлена группа племенных чистопородных пчелиных семей.
Среднегодовая прибавка медопродуктивности за счет селекции на одну пчелиную семью составила 11,2 кг, или 36 руб. (в ценах 1990 г.).
НИИ пчеловодства России была предложена схема и программа чистопородной селекции пчел, апробированная на Орловской и Башкирской опытных станциях пчеловодства, Приморском опорном пункте пчеловодства и в Грузинском НПО по пчеловодству.
Это объясняется, очевидно, особенностью серых горных кавказских пчел, заключающейся в их большей предприимчивости и способности эффективно использовать даже ничтожный медосбор. В этом случае значение большой силы пчелиных семей как бы нивелируется.
Сравнительно высокие значения коэффициентов корреляции были обнаружены у среднерусских пчел и помесей с их участием на Башкирской ОСП, Приморском ОПП и Чимкентском ОПП В этом случае на характер корреляций оказывала влияние такая породная особенность среднерусских пчел, как способность эффективно использовать лишь сильный медосбор.
Сходный с этим характер корреляций отмечен у пчелиных семей дальневосточной и краинской пород в местах их районирования (Приморский край и Чимкентская область Казахстана).
Сила корреляционных связей изменяется и в зависимости от условий сезона испытания, хотя и не в такой степени, как от происхождения пчелиных семей. В типичных для данной местности условиях следует ожидать, что различия будут несущественными. См.: Семьи-помеси под контроль! Старая история о наилучшей пчеле... Экологический гетерозис пчелиных семей и ошибки породного районирования. Породное районирование и лучшие пчелы для России. О гетерозисе.
ЗАКЛЮЧЕНИЕ
Пчелы среднерусской породы благодаря выработавшимся в процессе эволюции качествам — хорошей зимостойкости, устойчивости к ряду заболеваний, способности эффективно использовать короткий, но бурный медосбор, высокой плодовитости маток — незаменимы для разведения в центральных и северных зонах страны с их неблагоприятными условиями. К сожалению, в большинстве мест своего ареала среднерусские пчелы метизированы пчелами южных пород. Восстановление генофонда породы, размножение и селекционное улучшение— важная народнохозяйственная задача.
Генофонд среднерусской породы представлен рядом популяций, отличающихся морфологическими признаками. В процессе сравнительного испытания татарской, владимирской, новосибирской, уральской, вологодской и орловской популяций наиболее перспективной оказалась татарская, превысившая местных орловских пчел по среднесуточной яйценоскости маток на 17,9%, по медопродуктивности — на 25,7% и показавшая хорошую зимостойкость.
Исследования, проведенные за несколько десятилетий НИИ пчеловодства, позволили внести уточнения в методику определения чистопородности пчел и выделения популяций по экстерьерным признакам путем анализа их изменчивости. Определен объем выборки, необходимый для получения объективных данных по экстерьерным характеристикам внутрипородных группировок.
Изучение хозяйственно полезных признаков среднерусских пчел показало значительную фенотипическую изменчивость (особенно сильно варьируют показатели зимостойкости). Значительна степень генотипической изменчивости и наследуемости признаков, что дает основание рассчитывать на достаточную эффективность отбора. Однако эти параметры не остаются постоянными, они изменяются в процессе селекции и являются индикаторами его эффективности.
Установлены достоверные положительные фенотипические корреляции с медопродуктивностью пчел некоторых признаков: размеров крыла, тергита, величины кубитального индекса, зимостойкости, среднесуточной и максимальной яйценоскости маток, общей массы пчел перед медосбором.
Показано, что зимостойкость коррелирует с яйценоскостью маток при первом весеннем учете, со среднесуточной яйценоскостью за 36 дней до начала главного медосбора и с максимальной яйценоскостью. Все признаки, определяющие зимостойкость, коррелируют между собой, а также со степенью пораженности нозематозом, активностью каталазы ректальных желез, процентом сахарной добавки в зимних кормах. С максимальной яйценоскостью маток коррелирует их среднесуточная яйценоскость, а также их масса и размеры тела. Рассчитанные корреляции открывают широкие возможности для проведения косвенного отбора по сигнальным признакам.
Среднерусские пчелы отзывчивы на отбор. В среднем за 9 поколений превосходство над исходной популяцией составило по медопродуктивности и среднесуточной яйценоскости маток 60%, по максимальной яйценоскости — 40%. Наивысшие показатели среднесуточной яйценоскости маток и их максимальной яйценоскости были достигнуты в четвертом поколении.
Показана возможность использования высокого потенциала яйценоскости для производства на пасеках 100% пчелопакетов без существенного отрицательного влияния на медопродуктивность.
Поскольку при жестком отборе для селекционной работы уже к третьему поколению снижается генотипическое разнообразие признаков, усиливается их константность, уменьшается эффективность массового отбора, то наиболее перспективны для улучшения среднерусских пчел короткие линии. Для дальнейшего развития селективных признаков целесообразно использовать методы, повышающие их генотипическое разнообразие (кроссы линий, популяций).
Пчелы среднерусской породы отличаются хорошей сочетаемостью при скрещивании с другими породами, обеспечивающей получение эффекта гетерозиса как при простых двухпородных (генотип рабочих пчел), так и при сложных трехпородных скрещиваниях, затрагивающих и генотип маток. Так, среднее превосходство 8 вариантов сложных скрещиваний с использованием пчел среднерусской породы в четырех испытательных пунктах по яйценоскости составило 20%, а по медопродуктивности — 21% по сравнению с районированной породой. Положительный эффект создается за счет зимостойкости и яйценоскости маток, влияние которых распространяется и на другие признаки.
За 1976—1990 гг. в 30 областях районирования было реализовано более 80 тыс. неплодных и около 10 тыс. плодных маток отселекционированной линии № 44. Сплошной массив этой линии на Орловской опытной станции пчеловодства составил 1017 пчелиных семей.
На основе полученных материалов была организована и оформлена группа племенных чистопородных пчелиных семей.
Среднегодовая прибавка медопродуктивности за счет селекции на одну пчелиную семью составила 11,2 кг, или 36 руб. (в ценах 1990 г.).
НИИ пчеловодства России была предложена схема и программа чистопородной селекции пчел, апробированная на Орловской и Башкирской опытных станциях пчеловодства, Приморском опорном пункте пчеловодства и в Грузинском НПО по пчеловодству.
Зав. редакцией А.Н.Ивлев Художник Н.В.Медведева Художественный редактор В.В.Быков Технический редактор Л.П.Никитина Корректор В.В.Безымянская Слайды на обложке И.Н.Линича
В книге доктора сельскохозяйственных наук Н. И. Кривцова приведен богатейший материал селекционной работы с пчелами среднерусской породы, а также убедительно продемонстрировано преимущество этой породы для разведения в центральных и северных районах России.
Рассчитана как на ученых-селекционеров, так и на широкий круг пчеловодов-любителей.

Комментариев нет:
Отправить комментарий
Ваш комментарий должен соответствовать теме записи.